Submitted:
03 February 2025
Posted:
04 February 2025
You are already at the latest version
Abstract
Modern neuroscience understands the brain as a complex system whose functional properties are determined by the myriads of interactions of its cellular components. In the difficult route to obtain better understanding of this organ, mathematical tools have become ever more effective, and we are now able to simulate the dynamics of brain signals from digital representations that have high fidelity to experimental data. However, these simulations are generated by inherently simple neuronal models that contain assumptions that, by necessity, can be at times far removed from the underlying neurobiology. Here we develop an integrated view of brain dynamics that describes how the neurovascular units — made of neurons, capillaries and glia — collectively determine the development of the functional properties of brain neuronal populations and maintain their homeostatic control. These interactions are instructed by genetic programmes modulated by local environmental conditions. These complexities can be integrated into generative models and supply helpful analytics able to link the observable data to neurobiological entities if detailed brain mappings of molecular and cellular components are available. We further highlight how the development of these generative models could be relevant to the understanding of psychiatric conditions.
Keywords:
Brain
; dynamics
; metastability
; multi-stability
; fractality
; development
; homeostasis
; excitation
; inhibition
; astrocyte
; microglia
; complexity
; mathematical modelling
; abductive inference
1. Premise
Modern neuroscience understands the brain as a complex organ—an ensemble of billions of units that receive sensory signals from the environment and produce adaptive behavioural outputs as the result of a myriad of cellular computations taking place at various spatial and temporal scales (Barabasi and others 2023; BrynildsenRajanHenderson and Bassett 2023; Turkheimer and others 2022). However, scientific narratives that embrace this inherent complexity face a peculiar challenge, as efforts to build up from a comprehensive understanding of the biology and physiology of its cellular components often happen to need additional elements (e.g. computational models of particular subsystems) to properly account for higher levels of the organ, as scaling up from cells to populations and to systems is highly non-trivial (BassettZurn and Gold 2018; FornitoZalesky and Breakspear 2015; PulvermullerTomaselloHenningsen-Schomers and Wennekers 2021). Fortunately, important insights about how the brain operates at mesoscopic and macroscopic levels have been recently gained by analysing its functioning as a dynamic system (Brembs 2021). In this framework, the four concepts of connectivity, computation, criticality and coherence—the 4C's—have been widely used to characterise brain’s functionality in terms of its workings as a network, its capability to process information, its ability to switch rapidly across multiple states, and its capacity of transferring signals through dynamic modes of synchrony (Hancock and others 2022; LuppiRosasMedianoMenon and Stamatakis 2024; Sporns 2022). These research efforts have led to the hypothesis that healthy brain function depends on the balancing of two tendencies: stable integration between brain areas for effective coordinated functioning, and segregated processing within these same neuronal subsystems to utilize their functional specialisation (LordStevnerDeco and Kringelbach 2017; TononiEdelman and Sporns 1998). This unique signature has often been described as “metastability”.
In previous work we have reviewed the theoretical foundations and the empirical observations that have brought metastability to the fore of the Neurosciences (Hancock and others 2024). Complementing this work, here we investigate the biological underpinnings of such dynamic signature — which, so far, have been lacking a consistent, unified account. Indeed, not only metastability may be achieved via different mechanisms of varying levels of abstraction but, as we will see, even computational models with biological plausibility can be heavily dependent on parameter values and initial conditions to achieve a dynamical behaviour that successfully matches the observed brain dynamics (Castaldo and others 2023; Ramezanian-Panahi and others 2022; TouboulWendlingChauvel and Faugeras 2011; van Vreeswijk and Sompolinsky 1996). These facts lead to a natural question: how does the brain attains and subsequently stays into metastable regimes?
Here we propose that metastability is supported by two key properties, the small-worldness of structural and functional elements and excitation-inhibition balance. Crucially, we argue that these properties can only be properly understood when put together within the neurogliovascular system. We discuss the limited extent to which features of the neurogliovascular system are accounted for by current computational models, and avenues to improve this in the future. We also explore the potential role of neurogliovascular features on psychiatric conditions, and their potential to improve diagnosis and treatment.
2. Preliminaries
When investigating the origins of metastability, a natural approach is to study computational models of brain activity, starting with basic simulations of individual neurons and then moving up-scale to neuronal populations and networks. In this process, because of the limited number of parameters one can include into a computational model before it becomes intractable, details are progressively lost and new macroscopic features may be introduced; the construction by feature inclusion and exclusion is the actual mapping of the neurobiology into the model and its accuracy determines the face-validity of the results of the simulations but also of the analytics that can applied to the model when used with measurements (Ramezanian-Panahi and others 2022).
As between-neuron communication seems to be the principal determinant of brain dynamics, a good starting point for a computational model of the brain may be the generation of action potentials. The mechanism of this flow of ions was first explained by the influential Hodgkin-Huxley equations where the electrical current of the equivalent circuit is described by four differential equations that incorporate membrane capacity and the gating variables of the channels (Hodgkin and Huxley 1990). However, when the same model needs to be scaled up to population level, the circuitry needs to be simplified as multiple neuronal types need to be considered; moreover, the population structure must also be defined. For example, Izhikevic and Edelman’s (Izhikevich and Edelman 2008) whole cortex model used up to 22 different pyramidal and inhibitory interneurons, but adopted a simplified mono-equation dynamic that only modelled firing rates (as opposed to ion currents). This model can demonstrate several emergent phenomena such as self-sustained spontaneous activity, chaotic dynamics, and avalanches, alongside oscillatory activities like those in the human brain. The model is, however, very sensitive to the initial parameter settings and to the structural distribution of neuronal populations that, in this realization, was obtained from diffusion tensor images of adult subjects (please see next section) (Ramezanian-Panahi and others 2022). Hence this modelling approach is faced with two fundamental questions: how does the brain “get” into those initial conditions (e.g. how homeostasis is controlled), and what are the determinants of brain morphology. We explore these questions in the sequel.
2.1. Populations and Networks
Contemporary neuroscience understands brain function in health and development, ageing and disease, as emerging from widely distributed interactions of its units within and between spatial scales (cells, populations, anatomical regions etc) that may be conceptualised as networks (Papo and Buldú 2024; Rubinov 2024; Turkheimer and others 2022). Following this perspective, a natural question is to inquire about how these networks give rise to metastability, and which are their key properties in allowing this.
Simulation studies have convincingly demonstrated that the emergence of metastable dynamics is contingent on the pattern of interactions between the modules of a system (CabralHuguesSporns and Deco 2011; Friston 1997; PopeFukushimaBetzel and Sporns 2021; Shanahan 2010; Strogatz 2001). In particular, dynamic patterns of functional connectivity consistent with metastable dynamics have been observed to emerge when coupling (structural and functional) has “small-world” properties. Small-world networks (Watts and Strogatz 1998) are sets of nodes, or vertices, that are well connected such that neighbouring elements form highly connected clusters. These clusters are linked to the other distant ones by a smaller number of longer links so that the number of hops between any two of the vertices in the network is small (e.g. they have short-path lengths). Note that small-world networks may emerge from underlying self-similar (e.g. fractal) morphology at finer scales (BassettMeyer-LindenbergAchardDuke and Bullmore 2006). A number of previous investigations have demonstrated self-similar properties of neuronal dendritic branches which hold across a number of scales for each measurement(Caserta and others 1995; Kim and others 2011; Milosevic and Ristanovic 2006; MorigiwaTauchi and Fukuda 1989; SmithLange and Marks 1996; TakedaIshikawaOhtomoKobayashi and Matsuoka 1992; Wearne and others 2005).
Given the above, networks of anatomical connections derived from the human connectome data, that indeed possess small-world properties, have been incorporated within computational simulations of large-scale neural dynamics to demonstrate metastable properties of brain function (CabralHuguesSporns and Deco 2011; DecoJirsaMcIntoshSporns and Kotter 2009; HellyerScottShanahanSharp and Leech 2015; Honey and others 2009; OferShefi and Yaari 2017; Páscoa dos Santos and Verschure 2024). Empirical studies have then demonstrated that disruption to small-world brain networks is associated with neurodevelopmental and neurodegenerative disorders as well as brain injury (HellyerScottShanahanSharp and Leech 2015; Liu and others 2008; Pandit and others 2013; StamJonesNolteBreakspear and Scheltens 2007; Vasa and others 2015).
However, little is still known about the mechanisms driving such specific morphological architecture. Models put forward so far mirror the economy principle expressed by Ramón Cajal more than a century ago who proposed that neuronal morphology optimizes wiring (metabolic) costs (Ramón y Cajal 1899); the underlying mechanism of network optimization has been characterized as evolutionary, hence stemming from genetic control either generalized across the brain or adapting to the local cellular composition (AkarcaVértesBullmore and Astle 2021; Arnatkeviciute and others 2021; Bassett and Bullmore 2017; Bullmore and Sporns 2012; Libby and others 2017; MuldoonBridgeford and Bassett 2016; Oldham and others 2022; Vértes and others 2012).
We propose here that the assumption of complete genetic control of the development of small-world neural networks is not very plausible (Bremermann 1963; DuclosHendrikse and Jamniczky 2019). Indeed, the number of genes in the genome correlates almost linearly with the number of cell types in the various organisms, but it is far too low to code for the regulation of the interactions across all their elements (Kauffman 1991). Hence, a more likely assumption is that genetic programs may be activated or suppressed by local interactions, e.g. cell-to-cell information exchanges via epigenetic, hormonal, metabolic, chemical and electrical mechanisms that ultimately control cellular differentiation and biological function; this interconnected set of relation is generally referred to as the “physiome” and is described in detail in Section 3 (Bassingthwaighte 1992; DuclosHendrikse and Jamniczky 2019; Hunter and Borg 2003; Lindenmayer 1968).
2.2. Excitation-Inhibition Balance
A second key aspect of the emergence of brain dynamics is the combination of small-world networks with an appropriate balance between excitatory and inhibitory activities within the neuronal populations (Castaldo and others 2023; PoilHardstoneMansvelder and Linkenkaer-Hansen 2012; van Vreeswijk and Sompolinsky 1996), which we revise now.
Cellular units made by a pair of glutamatergic pyramidal (e.g. excitatory) and GABAergic (e.g. inhibitory) neurons constitute the canonical generators of brain signals (TurkheimerLeechExpertLord and Vernon 2015). The inhibitory action dampens spike-trains and morphs them into oscillations with frequency dependent on the feedback delay (Buzsaki and Wang 2012) (Figure 1).
For example, parvalbumin basket cells are activated by the excitatory signal transmitted by the paired pyramidal neurons and feedback negatively on the nucleus of these units with a delay of ~20 msec generating waves with gamma power of 50Hz (Borgers and Kopell 2003; Wang and Buzsaki 1996 ). Besides feeding back to the neurons they are coupled with, the same GABA interneurons also reach out to the inner part of the neuronal columns or project to the surrounding columns dampening the activities therein so that the traveling oscillations become 3-dimensional wave fronts (HelmstaedterSakmann and Feldmeyer 2009; TurkheimerLeechExpertLord and Vernon 2015). These oscillatory fronts are the binding mould the allows the synchronization of local neuronal ensembles but also, as they travel through distances in the brain, combine to generate slower waves that facilitate spatial synchronization across larger sections of the cortex (TurkheimerLeechExpertLord and Vernon 2015).
The basic cellular mechanisms described above support the use of mathematical oscillators as realistic models of brain functional units; these can be used at any scale, from the cellular to the regional to the population (Breakspear 2017; Strogatz 2001). Hence, one key aspect of the maintenance and propagation of signals in brain networks is the tight balance of excitation and inhibition within neuronal populations across scales. For example, in sensory cortical neurons increase in synaptic excitatory activity is followed closely by a concomitant increase in inhibitory conductance (Isaacson and Scanziani 2011; WuTao and Zhang 2011). A tight balance between synaptic excitation and inhibition is also observed during spontaneous network oscillations at rest (Isaacson and Scanziani 2011; Okun and Lampl 2008; ShuHasenstaub and McCormick 2003).
Excitation-inhibition heterogeneity across the brain and its apportioning of neuronal excitatory and inhibitory populations can be included into generative models (Deco and others 2021; Kong and others 2021; Zhang and others 2023). In these models, the tight balance between excitation and inhibition across the whole synaptic functional repertoire is generally achieved by adding to the model a plasticity component, e.g. the possibility of changes in parameters due to the interactions amongst inhibitory cell groupings or interactions between pyramidal and interneuronal clusters (Agnes and Vogels 2024; D'Amour J and Froemke 2015; Kirchner and Gjorgjieva 2021; Litwin-Kumar and Doiron 2014; MapelliGandolfiVilellaZoli and Bigiani 2016; VogelsSprekelerZenkeClopath and Gerstner 2011). These models have been able to demonstrate self-tuning mechanisms that, on one hand, keep synaptic connections in metastable dynamics and, at the same time, allow sufficient plasticity to remap sensory representations of new sets of stimuli (PoilHardstoneMansvelder and Linkenkaer-Hansen 2012; van Vreeswijk and Sompolinsky 1996; Yang and La Camera 2024).
However these attempts to biological fidelity are achieved by rules that are complex to implement as they are dependent on initial conditions, such as an appropriate ratio of GABA vs Glutamatergic neurons, and on the hand-tuning of model parameters; importantly, with a few exceptions (Agnes and Vogels 2024; Kirchner and Gjorgjieva 2021), they do not necessarily reflect cellular specific mechanisms.
2.3. From Small-World and E/I Balanced Networks to Metastability
The material above has reviewed two key aspects of the generative models used in the simulation of brain dynamics — that is, the structure of the networks (e.g. small worlds) and their homeostatic control (e.g. E/I balance). In this section we demonstrate how these two properties are important determine the transient states of dynamic connectivity that we have previously defined as metastability.
In order to elicit modularity, let us first note that the network needs some form of modularity. To see this, let us consider a toy example with three nodes — denoted by A, B, C — generating oscillatory activity. Let’s assume that B is the “hub” node, being connected to A and C, while A and C are not directly connected but, upon some general conditions, their connection to B allows a in-phase quick synchronization (Gollo and Breakspear 2014; Sporns and Kötter 2004). Under these conditions, the addition of any additional input into nodes A and C can easily “frustrate” the existing state of synchrony, so that the nodes A and C may then proceed to establish novel synchrony states using another hub node as pivot. This dynamic state of affair can be repeated across space and across scales wherever these motifs are implemented in a modular structure comprising a hub and 2 communities (Gollo and Breakspear 2014; Sporns and Kötter 2004). The resulting synchronized states equilibrate at frequencies that depend on the distances and the oscillatory states of the units of the motif (Gollo and Breakspear 2014; Sporns and Kötter 2004). This supports the idea that a small-world structure with distinctive neuronal hubs across scales may be suitable to induce metastable states to be formed across the swath of brain space.
At the same time, for the motifs to be reliably repeated across space and time, the neuronal masses should be able to maintain their oscillatory activity not withstanding a wide envelope of potential synaptic inputs as well as plastic adaptations necessary to brain development and function. Theoretical work has determined that, to maintain such robustness, a stable excitation-inhibition ratio based on strong excitatory and inhibitory currents internal to the oscillatory unit and stronger than any external input may be required (Páscoa dos Santos and Verschure 2024; van Vreeswijk and Sompolinsky 1996; van Vreeswijk and Sompolinsky 1998).
3. The Complexity of the Determinants of Brain Dynamics
Building on the considerations outlined above, here we posit that to build generative models that properly account for the brain’s biology we may need to consider the whole cellular repertoire of the tissue. This view poses substantial challenges, as each simulation is limited by the limited number of parameters that simulations can be consider at any one time. Accordingly, it is crucial to keep in mind that neural models bring with them a set of conditions related to non-neuronal determinants — which holds true even when neuronal only models are considered. In the sequel, we provide evidence that:
- The development of small-world neural networks can be understood in the context of their interactions with the development of vascular networks.
- Homeostasis, and specifically a tight E/I balance, relies heavily on the role of glia, and particularly astrocytes.
To advance the argument, we first briefly review the main neurobiological phases and determinants of brain development and particularly of neural and vascular networks, their mutual connections, as well as their interactions with glia. After this is done, next section highlights the potential relevance of these aspects to psychiatric.
3.1. The Development of Neural Networks
The formation of branched structures that ultimately shape neuronal networks relies on processes of arborization and polarization that take place during neurogenesis in the development of each neuron (GuidolinFede and Tortorella 2019). As reviewed below, such processes rely on the concerted switching on-and-off of genetic programs based on other cellular interactions.
Brain development proceeds in overlapping phases: the making of the brain cells, cell migration, axons and dendrites growth, synaptic development and linkage with other neuronal cells, synaptic maturation and pruning, glia-genesis and myelination (ShonkoffPhillips and National Research Council (U.S.). Committee on Integrating the Science of Early Childhood Development. 2000) (Figure 2).
The brain and spinal cord arise from a set of cells on the dorsal part (e.g. neural plate) of the developing embryo. The dividing cells proliferate on each side of the plate along its length and their folding forms the neural tube. In the tube, three contained enlargements are formed from which the various parts of the brain are generated; the anterior portion gives rise to the cerebral hemispheres, the middle section to the midbrain, the posterior to the brain stem, the cerebellum and the spinal cord and peripheral nerves. Under the control of regulatory genes, the neural stem/progenitor cells (NSCs/NPCs) start a first differentiation stage mostly converting into radial glial cells (RGCs); these cells are polarized and stretch radially from the early ventricles to the pia becoming the pillars of the expanding cortex. This is the scaffolding that guides the next phase when neurons travel to their appropriate layer in the neural columns (ShonkoffPhillips and National Research Council (U.S.). Committee on Integrating the Science of Early Childhood Development. 2000). This process stretches until the 15th gestational week after which astrocytes and oligodendrocytes commence their proliferation from RGCs and migration to support the developing neural columns. Note that, differently from the other glia, microglial cells appear very early in gestation (from week 8) and then distribute across the developing cortex (Menassa and Gomez-Nicola 2018).
Once the nerve cells are formed and finish migrating, they rapidly extend axons, often over relatively long distances, as well as dendrites and begin to form connections with other nerve cells; these connections are called synapses. This process continues well into the childhood years. There is evidence in many parts of the nervous system that the stability and strength of these synapses are largely determined by synaptic activity, hence are dependent on environmental stimuli. As nerve cells form axons, accompanying oligodendrocytes wrap them with an insulating coating, myelin, that increases conduction velocity.
Axon guidance is a complex process whereas guidance receptors expressed at the tips of motile axons interact with guidance cues; these cues present with both positive (attraction) and negative (repulsion) feedback systems that are locally switched on and off in temporal sequelae along the path to avoid the formation of loops (ZangChaudhari and Bashaw 2021). Environmental clues also drive the arborization of axons, the process that extend multiple branches from the tip end to form terminal arbores (Kalil and Dent 2014). The most important of these interactions are those with the vascular network.
The brain is one of the most energy consuming organs whose metabolism is mostly reliant on oxidative metabolism and holds minimal glycogen reserve, hence it requires a constant supply of oxygen and glucose (LordExpertHuckins and Turkheimer 2013; Oz and others 2007). The distribution of synapses, where most of the energy budget is spent with 50% used on post-synaptic glutamate receptors, must then spatially match the blood vessel area that supplies glucose and oxygen (HowarthGleeson and Attwell 2012; HowarthPeppiatt-Wildman and Attwell 2010).
The close interaction between neural and vascular networks in the brain is well documented and has even gained a specific label by being called the “neurovascular unit” (Eichmann and Thomas 2013; Iadecola 2017).
Hence, we propose that the small-world brain morphology of neural networks is based on the small-world morphology of the neurovascular unit. It is relevant to note that other organs that have very high metabolic demand, if not higher than the brain, such as such as kidneys, heart and liver, have all been shown to possess branching morphology and that this is not the case for other organs with lower metabolic needs (e.g. adipose tissue and skeletal muscle) (CapturKarperienHughesFrancis and Moon 2017; Flanagan and others 2021; Smith and Trachtenberg 2007; Wang and others 2010). We develop this argument further in the following section.
3.2. The Development and Cross-Talk of Vascular Networks
Contrary to neural networks, the mechanistic bases of small-world development and morphology are well understood and modelled as fractal biology when it comes to the vascular system (Bassingthwaighte 1992). The morphogenesis of vascular networks is based on the proliferative activity of equipotent tips that branch out to respond to metabolic demands, e.g. hypoxia, and become proliferatively inactive when the latter are quenched and/or they enter an already oxygenated territory; whilst expanding, they also supress the sprouting of neighbouring tips (Hannezo and others 2017). Hence, vascular branched structures develop as a self-organized process, rely on simple local rules, and do not require a deterministic sequence of genetically programmed events (Cardy and Tauber 1996; Hannezo and others 2017).
Specifically, the process of vascular branching is based on two elegant feedback systems (Figure 4). During development, tissue expansion causes hypoxic conditions that are sensed by the local astrocytic population that releases the vascular endothelial growth factor (VEGF). VEGF release stimulates the sprouting of endothelial cells that expand the vascular network and bring oxygenated blood until VEGF expression is switched off (Gerhardt and others 2003). For such branching system to work, only a few endothelial cells need to be selected as tips while the others trail these and form tubes; this system is controlled via a form of lateral inhibition that blocks sprouting from adjacent cells via the Notch signalling pathway. (Roca and Adams 2007).
Importantly for what is purported here, there is a close cross-talk between the vascular and neural networks that share signals through known axon and blood vessel guidance molecules: the Neuropilin receptors and their Semaphorin and VEGF ligands, and Netrins and Slits ligands and their receptors (Carmeliet and Tessier-Lavigne 2005; Eichmann and Thomas 2013; Iadecola 2017; Jones and Li 2007). A few models have been proposed to demonstrate the development of brain vasculature (Ii and others 2020; KarchNeumannNeumann and Schreiner 1999; PeyrounetteDavitQuintard and Lorthois 2018; ShipleySmithSweeneyPries and Secomb 2020); Kumar and colleagues have proposed the first synthetic formulation of the interaction between growing vascular and neuronal networks (KumarMenonGayathri and Chakravarthy 2022).
3.3. The Control of Excitation-Inhibition Ratio Balance
Approximately one in six neurons in the adult neocortex are inhibitory GABAergic interneurons, and this ratio is relatively stable across development but not across layers — with higher densities observed in layers 2 and 5 (Beaulieu 1993; Goulas and others 2021; Meyer and others 2011; SaharaYanagawaO'Leary and Stevens 2012). GABA interneurons are a highly heterogeneous group of neurons with diverse morphologies, biochemical and physiological properties (LimMiLlorca and Marin 2018). These features are acquired during development through the implementation of specific transcriptional programs that are intrinsically encoded and switched by local cues or driven by local interactions with the microenvironment (LimMiLlorca and Marin 2018 ). In particular, the laminar distribution, phenotype, connectivity and even survival of interneurons are determined by signals provided by pyramidal cells (HevnerDazaEnglundKohtz and Fink 2004; Lodato and others 2011; Miyoshi and others 2010; Pla and others 2018; Southwell and others 2012; Ye and others 2015). The maturation of interneurons and of their connections to pyramidal neurons drives the early appearance of local networks and gamma oscillations (gestation week 24) until full maturation of brain dynamics up to two years from birth (WuKourdougli and Portera-Cailliau 2024).
Interneurons have well established role in inducing long-term plasticity at excitatory synapses (Wigstrom and Gustafsson 1985); at the same time, as the result of interaction between excitatory and inhibitory synapses, excitatory transmission modulates inhibitory synaptic plasticity (Belan and Kostyuk 2002). These cross-modulatory effects are well captured by a few of the mathematical models proposed (PulvermullerTomaselloHenningsen-Schomers and Wennekers 2021).
However, current mathematical models do not account for glial cells (Fields and others 2014; Ramezanian-Panahi and others 2022). Glial cells are the most abundant (90%) cells in the human brain and are tightly integrated into neural networks. As already reviewed in previous sections, they have a key role in the development of vascular and neural networks and, in the mature brain, they are responsible for homeostatic control. They sense and shuffle major ions, remove and catabolize neurotransmitters and supply neurons with neurotransmitter precursors; crucially they maintain metabolic homeostasis by metabolizing glucose and supplying neurons with energy substrates, mostly lactate, and by synthesizing glycogen (Verkhratsky and Nedergaard 2018). With their perisynaptic sheaths, astrocytes enwrap the whole synapses and regulate synaptic maturation, maintenance, and extinction (Verkhratsky and Nedergaard 2014).
Moreover, astrocytes use their large array of transporters to control the dynamics of neurotransmitter concentration in the synaptic cleft and modulate a wide array of synaptic responses (Marcaggi and Attwell 2004). For example, astroglial release of glutamate may results in both inhibition or potentiation of evoked and spontaneous excitatory or inhibitory postsynaptic currents as well as modulation of long-term potentiation or long-term depression (AraqueParpuraSanzgiri and Haydon 1998; Han and others 2012; Jourdain and others 2007; KangJiangGoldman and Nedergaard 1998; LiuXuArcuinoKang and Nedergaard 2004; Min and Nevian 2012; Navarrete and others 2012; SantelloBezzi and Volterra 2011; SantelloCali and Bezzi 2012). Conversely astroglial GABA transporters have a key role in controlling concentration of GABA in the synaptic cleft hence control GABA tone (Kersante and others 2013).
4. The Relevance of the Neurogliovascular Unit to Psychiatric Disorders
The previous section has summarised how proper cerebrovascular development and neurogliovascular unit assembly are essential for brain growth and function, during development and throughout life, ensuring the continuous supply of nutrients and oxygen. This supports the view that the inclusion into computational models of components of the neurogliovascular unit is relevant to contemporary neuropsychiatry — a view that we explore now.
Unsurprisingly, abnormalities in these vascular and glial compartments have been involved in the pathophysiology of psychiatric disorders. (Clark-Raymond and Halaris 2013; CollignonDion-AlbertMénard and Coelho-Santos 2024; Lee and Kim 2012; PalmerWillhoite and Gage 2000). VEGF plays a role not only in cerebrovascular development but also in neurogenesis in the adult brain and its dysregulation has been consistently associated with major depressive disorder (Clark-Raymond and Halaris 2013; CollignonDion-AlbertMénard and Coelho-Santos 2024; Lee and Kim 2012; Nunes and others 2022; PalmerWillhoite and Gage 2000; Xie and others 2017). Signalling pathways that we have reviewed above as key to microvascular development have also been linked to developmental disorders, schizophrenia, bipolar disorder and autism (Hoseth and others 2018; WangYu and Li 2023; YangGuanSheng and Gu 2024). Microvascular abnormalities measured by retinal imaging in fact are a consistent finding in patients with schizophrenia (KatselRoussosPletnikov and Haroutunian 2017; Meier and others 2013; WangYu and Li 2023).
However, morphological information about abnormalities of the brain microvasculature in psychiatric cohorts is scant. In particular, total cerebral blood volume has been reported as below normal levels in patients with schizophrenia (Brambilla and others 2007) and this seems to be the result also of underdeveloped small arterial (pial) and arteriolar vessels (Hua and others 2017). It is worth noting that MRI methods to quantify the density of both arterial and venous micro-vessels are available (Balchandani and Naidich 2015; Hua and others 2011).
The relevance of excitation-inhibition imbalance in psychiatric disorders has been widely documented with findings in clinical cohorts with schizophrenia, autism spectrum disorder as well as mood disorders (see (TurkheimerLeechExpertLord and Vernon 2015) for a review) and the estimation of local excitation-inhibition balance is now an important effort in the development of analytics for brain functional imaging data (Colombo and others 2019; Deco and others 2014; GaoPeterson and Voytek 2017; Páscoa dos Santos and Verschure 2024; Podvalny and others 2015; Waschke and others 2021; Zhang and others 2023). Direct in-vivo quantification of glutamate (Glu), and γ-aminobutyric acid (GABA) steady-state concentrations can be achieved using Magnetic Resonance Spectroscopy (MRS) (Rae 2014). However, it is still challenging to apportion the concentrations of these neurotransmitters to the underlying cellular mechanisms, e.g. neuronal or glial (Lea-CarnallEl-DeredyStaggWilliams and Trujillo-Barreto 2023). Initial approaches included the use of metabolic activity in astrocytes as a proxy of cycling Glu given its high metabolic costs (RothmanBeharHyder and Shulman 2003); only very recently MRS has been used to quantify time-resolved metabolic responses to stimuli on much shorter timescales of seconds to minutes, so-called functional MRS (JelenKingMullins and Stone 2018), and these signals have allowed the attribution of Glu and GABA concentrations to cellular compartments (Lea-CarnallEl-DeredyStaggWilliams and Trujillo-Barreto 2023).
5. Conclusions
In this manuscript we have provided a critical view on how mathematical models that are used to simulate neural dynamic rely on a variety of properties that are directly tight with various aspects of brain biology. These models are bounded to provided simplified representations that may have difficulties in properly capturing the rich complexity of the cellular and molecular processes taking place underneath. Concretely, so far they have mostly failed to include non-neuronal cells, glia and the vascular system, that are important determinants of brain morphology and function. Importantly, most approaches miss the general principle that the brain’s homeostasis is achieved by the set of interactions amongst cell types (e.g. the physiome (Hunter and Borg 2003)), and that cell differentiation during development is itself a locally controlled process (Rowitch and Kriegstein 2010).
Given the lack of direct measurements in clinical populations, it is not surprising that the relevance of the neurogliovascular unit in neuropsychiatric disorders has not been matched by adequate computational modelling efforts. If direct measurements of these non-neuronal components and comprehensive molecular and cellular mapping will be made available, these components could allow us to further constraint the ranges of the model’s parameters. This, in turn, has the potential to improve our ability to properly interpret the results obtained from these models and contribute to the incremental, gradual process of discovery of the biology of brain in health and disease. This process is made more effective by the recent availability of ever more detailed and systematic information on the brain at any scale (e.g. molecular, cellular and behavioural) (Calzavarini and Cevolani 2022; Lawn and others 2023; Markello and others 2022; YarkoniPoldrackNicholsVan Essen and Wager 2011). For example, substantial improvement in model fit to functional MRI data has been observed by the inclusion of intracortical myelin density to constrain model by parameterizing synaptic strength (Deco and others 2021; Demirtaş and others 2019). Recently, the first molecular atlas of the cells that make-up the human vasculature has been reported and has clearly resolved molecular components that are key to the pathophysiology of neurodegenerative disorders (Yang and others 2022).
All the above warrants great promise to the use of physical models and mathematical tools to study a wide range of topics with high potential translational value including, for example, the role of the vascular component and/or excitation-inhibition balance for several disorders affecting higher cognitive functions such as major depression, autism, and schizophrenia (AzmitiaSaccomanoAlzoobaeeBoldrini and Whitaker-Azmitia 2016; Hanson and Gottesman 2005; Hollestein and others 2023; Liu and others 2021). We hope this work may inspire more investigations on the rich interplay between neural, glial, and vascular activity, which we regard as critical to foster a greater understanding of the arguably most fascinating and mysterious organ, the brain.
Acknowledgments
FET is partly funded by the NIHR Maudsley Biomedical Research Centre at South London and Maudsley NHS Foundation Trust and King’s College London.
References
- Agnes, E.J.; Vogels, T.P. Co-dependent excitatory and inhibitory plasticity accounts for quick, stable and long-lasting memories in biological networks. Nat. Neurosci. 2024, 27, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Akarca, D.; Vértes, P.E.; Bullmore, E.T.; Astle, D.E. A generative network model of neurodevelopmental diversity in structural brain organization. Nat Commun 2021, 12, 4216. [Google Scholar] [PubMed]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur. J. Neurosci. 1998, 10, 2129–2142. [Google Scholar] [CrossRef] [PubMed]
- Arnatkeviciute, A.; Fulcher, B.D.; Oldham, S.; Tiego, J.; Paquola, C.; Gerring, Z.; Aquino, K.; Hawi, Z.; Johnson, B.; Ball, G.; et al. Genetic influences on hub connectivity of the human connectome. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Azmitia, E.C.; Saccomano, Z.T.; Alzoobaee, M.F.; Boldrini, M.; Whitaker-Azmitia, P.M. Persistent Angiogenesis in the Autism Brain: An Immunocytochemical Study of Postmortem Cortex, Brainstem and Cerebellum. J. Autism Dev. Disord. 2015, 46, 1307–1318. [Google Scholar] [CrossRef]
- Balchandani, P.; Naidich, T.P. Ultra-High-Field MR Neuroimaging. AJNR Am J Neuroradiol 2015, 36, 1204–15. [Google Scholar] [CrossRef]
- Barabási, D.L.; Bianconi, G.; Bullmore, E.; Burgess, M.; Chung, S.; Eliassi-Rad, T.; George, D.; Kovács, I.A.; Makse, H.; Nichols, T.E.; et al. Neuroscience Needs Network Science. J. Neurosci. 2023, 43, 5989–5995. [Google Scholar] [CrossRef]
- Bassett, D.S.; Bullmore, E.T. Small-World Brain Networks Revisited. Neuroscientist 2017, 23, 499–516. [Google Scholar]
- Bassett, D.S.; Meyer-Lindenberg, A.; Achard, S.; Duke, T.; Bullmore, E. Adaptive reconfiguration of fractal small-world human brain functional networks. Proc. Natl. Acad. Sci. 2006, 103, 19518–19523. [Google Scholar] [CrossRef]
- Bassett, D.S.; Zurn, P.; Gold, J.I. On the nature and use of models in network neuroscience. Nat. Rev. Neurosci. 2018, 19, 566–578. [Google Scholar] [CrossRef]
- Bassingthwaighte, J.B. Fractal Vascular Growth Patterns. Acta Stereol 1992, 11 (Suppl. 1), 305–319. [Google Scholar] [PubMed]
- Beaulieu, C. Numerical data on neocortical neurons in adult rat, with special reference to the GABA population. Brain Res. 1993, 609, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Belan, P.; Kostyuk, P. Glutamate-receptor-induced modulation of GABAergic synaptic transmission in the hippocampus. Pfl?gers Arch. Eur. J. Physiol. 2002, 444, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Börgers, C.; Kopell, N. Synchronization in Networks of Excitatory and Inhibitory Neurons with Sparse, Random Connectivity. Neural Comput. 2003, 15, 509–538. [Google Scholar] [CrossRef]
- Brambilla, P.; Cerini, R.; Fabene, P.F.; Andreone, N.; Rambaldelli, G.; Farace, P.; Versace, A.; Perlini, C.; Pelizza, L.; Gasparini, A.; et al. Assessment of cerebral blood volume in schizophrenia: A magnetic resonance imaging study. J. Psychiatr. Res. 2007, 41, 502–510. [Google Scholar] [CrossRef]
- Breakspear, M. Dynamic models of large-scale brain activity. Nat. Neurosci. 2017, 20, 340–352. [Google Scholar] [CrossRef]
- Brembs, B. The brain as a dynamically active organ. Biochem Biophys Res Commun 2020, 564, 55–69. [Google Scholar] [CrossRef]
- Bremermann, H.J. Limits of Genetic Control. IEEE Transactions on Military Electronics 1963, MIL-7, 200–205. [Google Scholar] [CrossRef]
- Brynildsen, J.K.; Rajan, K.; Henderson, M.X.; Bassett, D.S. Network models to enhance the translational impact of cross-species studies. Nat. Rev. Neurosci. 2023, 24, 575–588. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef]
- Buzsáki, G.; Wang, X.-J. Mechanisms of Gamma Oscillations. Annu. Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.; Hugues, E.; Sporns, O.; Deco, G. Role of local network oscillations in resting-state functional connectivity. NeuroImage 2011, 57, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Calzavarini, F.; Cevolani, G. Abductive reasoning in cognitive neuroscience: weak and strong reverse inference. Synthese 2022, 200, 1–26. [Google Scholar] [CrossRef]
- Captur, G.; Karperien, A.L.; Hughes, A.D.; Francis, D.P.; Moon, J.C. The fractal heart — embracing mathematics in the cardiology clinic. Nat. Rev. Cardiol. 2016, 14, 56–64. [Google Scholar] [CrossRef]
- Cardy, J.; Täuber, U.C. Theory of Branching and Annihilating Random Walks. Phys. Rev. Lett. 1996, 77, 4780–4783. [Google Scholar] [CrossRef]
- Carmeliet, P.; Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature 2005, 436, 193–200. [Google Scholar] [CrossRef]
- Caserta, F.; Eldred, W.; Fernandez, E.; Hausman, R.; Stanford, L.; Bulderev, S.; Schwarzer, S.; Stanley, H. Determination of fractal dimension of physiologically characterized neurons in two and three dimensions. J. Neurosci. Methods 1995, 56, 133–144. [Google Scholar] [CrossRef]
- Castaldo, F.; dos Santos, F.P.; Timms, R.C.; Cabral, J.; Vohryzek, J.; Deco, G.; Woolrich, M.; Friston, K.; Verschure, P.; Litvak, V. Multi-modal and multi-model interrogation of large-scale functional brain networks. NeuroImage 2023, 277, 120236. [Google Scholar] [CrossRef]
- Clark-Raymond, A.; Halaris, A. VEGF and depression: A comprehensive assessment of clinical data. J. Psychiatr. Res. 2013, 47, 1080–1087. [Google Scholar] [CrossRef]
- Collignon, A.; Dion-Albert, L.; Ménard, C.; Coelho-Santos, V. Sex, hormones and cerebrovascular function: from development to disorder. Fluids Barriers CNS 2024, 21, 1–19. [Google Scholar] [CrossRef]
- Colombo, M.A.; Napolitani, M.; Boly, M.; Gosseries, O.; Casarotto, S.; Rosanova, M.; Brichant, J.-F.; Boveroux, P.; Rex, S.; Laureys, S.; et al. The spectral exponent of the resting EEG indexes the presence of consciousness during unresponsiveness induced by propofol, xenon, and ketamine. 2019, 189, 631–644. [Google Scholar] [CrossRef] [PubMed]
- D’amour, J.A.; Froemke, R.C. Inhibitory and Excitatory Spike-Timing-Dependent Plasticity in the Auditory Cortex. Neuron 2015, 86, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Deco G, Jirsa V, McIntosh AR, Sporns O, Kotter R. 2009. Key role of coupling, delay, and noise in resting brain fluctuations. Proc Natl Acad Sci U S A 2009, 106, 10302-7.
- Deco, G.; Kringelbach, M.L.; Arnatkeviciute, A.; Oldham, S.; Sabaroedin, K.; Rogasch, N.C.; Aquino, K.M.; Fornito, A. Dynamical consequences of regional heterogeneity in the brain’s transcriptional landscape. Sci. Adv. 2021, 7, eabf4752. [Google Scholar] [CrossRef]
- Deco, G.; Ponce-Alvarez, A.; Hagmann, P.; Romani, G.L.; Mantini, D.; Corbetta, M. How Local Excitation-Inhibition Ratio Impacts the Whole Brain Dynamics. J. Neurosci. 2014, 34, 7886–7898. [Google Scholar] [CrossRef]
- Demirtaş, M.; Burt, J.B.; Helmer, M.; Ji, J.L.; Adkinson, B.D.; Glasser, M.F.; Van Essen, D.C.; Sotiropoulos, S.N.; Anticevic, A.; Murray, J.D. Hierarchical Heterogeneity across Human Cortex Shapes Large-Scale Neural Dynamics. Neuron 2019, 101, 1181–1194.e13. [Google Scholar] [CrossRef]
- Duclos, K.K.; Hendrikse, J.L.; Jamniczky, H.A. Investigating the evolution and development of biological complexity under the framework of epigenetics. Evol. Dev. 2019, 21, 276–293. [Google Scholar] [CrossRef]
- Eichmann, A.; Thomas, J.-L. Molecular Parallels between Neural and Vascular Development. Cold Spring Harb. Perspect. Med. 2013, 3, a006551. [Google Scholar] [CrossRef]
- Fields, R.D.; Araque, A.; Johansen-Berg, H.; Lim, S.-S.; Lynch, G.; Nave, K.-A.; Nedergaard, M.; Perez, R.; Sejnowski, T.; Wake, H. Glial Biology in Learning and Cognition. Neurosci. 2013, 20, 426–431. [Google Scholar] [CrossRef]
- Flanagan, B.; McDaid, L.; Wade, J.J.; Toman, M.; Wong-Lin, K.; Harkin, J. A Computational Study of Astrocytic GABA Release at the Glutamatergic Synapse: EAAT-2 and GAT-3 Coupled Dynamics. Front. Cell. Neurosci. 2021, 15, 682460. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Breakspear, M. The connectomics of brain disorders. Nat. Rev. Neurosci. 2015, 16, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Transients, metastability, and neuronal dynamics. Neuroimage 1997, 5, 164–71. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Peterson, E.J.; Voytek, B. Inferring synaptic excitation/inhibition balance from field potentials. NeuroImage 2017, 158, 70–78. [Google Scholar] [CrossRef]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Gollo, L.L.; Breakspear, M. The frustrated brain: from dynamics on motifs to communities and networks. Philos. Trans. R. Soc. B: Biol. Sci. 2014, 369, 20130532. [Google Scholar] [CrossRef] [PubMed]
- Goulas, A.; Changeux, J.-P.; Wagstyl, K.; Amunts, K.; Palomero-Gallagher, N.; Hilgetag, C.C. The natural axis of transmitter receptor distribution in the human cerebral cortex. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Guidolin, D.; Fede, C.; Tortorella, C. Nerve cells developmental processes and the dynamic role of cytokine signaling. Int. J. Dev. Neurosci. 2018, 77, 3–17. [Google Scholar] [CrossRef]
- Han, J.; Kesner, P.; Metna-Laurent, M.; Duan, T.; Xu, L.; Georges, F.; Koehl, M.; Abrous, D.N.; Mendizabal-Zubiaga, J.; Grandes, P.; et al. Acute Cannabinoids Impair Working Memory through Astroglial CB1 Receptor Modulation of Hippocampal LTD. Cell 2012, 148, 1039–1050. [Google Scholar] [CrossRef]
- Hancock, F.; Rosas, F.E.; Luppi, A.I.; Zhang, M.; Mediano, P.A.M.; Cabral, J. Metastability demystified - the foundational past, the pragmatic present and the promising future. Nat Rev Neurosci. 2024. [CrossRef]
- Hancock, F.; Rosas, F.E.; Mediano, P.A.M.; Luppi, A.I.; Cabral, J.; Dipasquale, O. May the 4C's be with you: an overview of complexity-inspired frameworks for analysing resting-state neuroimaging data. J R Soc Interface 2022, 19, 20220214. [Google Scholar] [CrossRef]
- Hannezo, E.; Scheele, C.L.G.J.; Moad, M.; Drogo, N.; Heer, R.; Sampogna, R.V.; van Rheenen, J.; Simons, B.D. A Unifying Theory of Branching Morphogenesis. Cell 2017, 171, 242–255.e27. [Google Scholar] [CrossRef] [PubMed]
- Hanson, D.R.; I Gottesman, I. Theories of schizophrenia: a genetic-inflammatory-vascular synthesis. BMC Med Genet. 2005, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hellyer, P.J.; Scott, G.; Shanahan, M.; Sharp, D.J.; Leech, R. Cognitive Flexibility through Metastable Neural Dynamics Is Disrupted by Damage to the Structural Connectome. J. Neurosci. 2015, 35, 9050–9063. [Google Scholar] [CrossRef]
- Helmstaedter, M.; Sakmann, B.; Feldmeyer, D. Neuronal Correlates of Local, Lateral, and Translaminar Inhibition with Reference to Cortical Columns. Cereb. Cortex 2008, 19, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Hevner, R.; Daza, R.; Englund, C.; Kohtz, J.; Fink, A. Postnatal shifts of interneuron position in the neocortex of normal and reeler mice: evidence for inward radial migration. Neuroscience 2004, 124, 605–618. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. Bull Math Biol 1990, 52, 25–71; discussion 5. [Google Scholar] [CrossRef]
- Hollestein, V.; Poelmans, G.; Forde, N.J.; Beckmann, C.F.; Ecker, C.; Mann, C.; Schäfer, T.; Moessnang, C.; Baumeister, S.; Banaschewski, T.; et al. Excitatory/inhibitory imbalance in autism: the role of glutamate and GABA gene-sets in symptoms and cortical brain structure. Transl. Psychiatry 2023, 13, 1–9. [Google Scholar] [CrossRef]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting human resting-state functional connectivity from structural connectivity. Proc. Natl. Acad. Sci. 2009, 106, 2035–2040. [Google Scholar] [CrossRef]
- Hoseth, E.Z.; Krull, F.; Dieset, I.; Mørch, R.H.; Hope, S.; Gardsjord, E.S.; Steen, N.E.; Melle, I.; Brattbakk, H.-R.; Steen, V.M.; et al. Attenuated Notch signaling in schizophrenia and bipolar disorder. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Howarth, C.; Gleeson, P.; Attwell, D. Updated Energy Budgets for Neural Computation in the Neocortex and Cerebellum. J. Cereb. Blood Flow Metab. 2012, 32, 1222–1232. [Google Scholar] [CrossRef]
- Howarth, C.; Peppiatt-Wildman, C.M.; Attwell, D. The Energy Use Associated with Neural Computation in the Cerebellum. J. Cereb. Blood Flow Metab. 2009, 30, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Brandt, A.S.; Lee, S.; Blair, N.I.S.; Wu, Y.; Lui, S.; Patel, J.; Faria, A.V.; Lim, I.A.L.; Unschuld, P.G.; et al. Abnormal Grey Matter Arteriolar Cerebral Blood Volume in Schizophrenia Measured With 3D Inflow-Based Vascular-Space-Occupancy MRI at 7T. Schizophr. Bull. 2016, 43, 620–632. [Google Scholar] [CrossRef]
- Hua, J.; Qin, Q.; Donahue, M.J.; Zhou, J.; Pekar, J.J.; van Zijl, P.C. Inflow-based vascular-space-occupancy (iVASO) MRI. Magn Reson Med 2011, 66, 40–56. [Google Scholar] [PubMed]
- Hunter, P.J.; Borg, T.K. Integration from proteins to organs: the Physiome Project. Nat. Rev. Mol. Cell Biol. 2003, 4, 237–243. [Google Scholar] [CrossRef]
- Iadecola, C. The Neurovascular Unit Coming of Age: A Journey through Neurovascular Coupling in Health and Disease. Neuron 2017, 96, 17–42. [Google Scholar] [CrossRef]
- Ii, S.; Kitade, H.; Ishida, S.; Imai, Y.; Watanabe, Y.; Wada, S. Multiscale modeling of human cerebrovasculature: A hybrid approach using image-based geometry and a mathematical algorithm. PLOS Comput. Biol. 2020, 16, e1007943. [Google Scholar] [CrossRef]
- Isaacson, J.S.; Scanziani, M. How Inhibition Shapes Cortical Activity. Neuron 2011, 72, 231–243. [Google Scholar] [CrossRef]
- Izhikevich, E.M.; Edelman, G.M. Large-scale model of mammalian thalamocortical systems. Proc. Natl. Acad. Sci. 2008, 105, 3593–3598. [Google Scholar] [CrossRef]
- A Jelen, L.; King, S.; Mullins, P.G.; Stone, J.M. Beyond static measures: A review of functional magnetic resonance spectroscopy and its potential to investigate dynamic glutamatergic abnormalities in schizophrenia. J. Psychopharmacol. 2018, 32, 497–508. [Google Scholar] [CrossRef]
- A Jones, C.; Li, D.Y. Common cues regulate neural and vascular patterning. Curr. Opin. Genet. Dev. 2007, 17, 332–336. [Google Scholar] [CrossRef]
- Jourdain, P.; Bergersen, L.H.; Bhaukaurally, K.; Bezzi, P.; Santello, M.; Domercq, M.; Matute, C.; Tonello, F.; Gundersen, V.; Volterra, A. Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 2007, 10, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Kalil, K.; Dent, E.W. Branch management: mechanisms of axon branching in the developing vertebrate CNS. Nat. Rev. Neurosci. 2013, 15, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Jiang, L.; Goldman, S.A.; Nedergaard, M. Astrocyte-mediated potentiation of inhibitory synaptic transmission. Nat. Neurosci. 1998, 1, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Karch, R.; Neumann, F.; Neumann, M.; Schreiner, W. A three-dimensional model for arterial tree representation, generated by constrained constructive optimization. Comput. Biol. Med. 1999, 29, 19–38. [Google Scholar] [CrossRef]
- Katsel, P.; Roussos, P.; Pletnikov, M.; Haroutunian, V. Microvascular anomaly conditions in psychiatric disease. Schizophrenia – angiogenesis connection. Neurosci. Biobehav. Rev. 2017, 77, 327–339. [Google Scholar] [CrossRef]
- Kauffman, S.A. Antichaos and Adaptation. Sci. Am. 1991, 265, 78–84. [Google Scholar] [CrossRef]
- Kersanté, F.; Rowley, S.C.S.; Pavlov, I.; Gutièrrez-Mecinas, M.; Semyanov, A.; Reul, J.M.H.M.; Walker, M.C.; Linthorst, A.C.E. A functional role for both γ-aminobutyric acid (GABA) transporter-1 and GABA transporter-3 in the modulation of extracellular GABA and GABAergic tonic conductances in the rat hippocampus. J. Physiol. 2013, 591, 2429–2441. [Google Scholar] [CrossRef]
- Kim, J.; Kwon, N.; Chang, S.; Kim, K.-T.; Lee, D.; Kim, S.; Yun, S.J.; Hwang, D.; Kim, J.W.; Hwu, Y.; et al. Altered branching patterns of Purkinje cells in mouse model for cortical development disorder. Sci. Rep. 2011, 1, 122. [Google Scholar] [CrossRef]
- Kirchner, J.H.; Gjorgjieva, J. Emergence of local and global synaptic organization on cortical dendrites. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Kong, X.; Kong, R.; Orban, C.; Wang, P.; Zhang, S.; Anderson, K.; Holmes, A.; Murray, J.D.; Deco, G.; Heuvel, M.v.D.; et al. Sensory-motor cortices shape functional connectivity dynamics in the human brain. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Kumar, B.S.; Menon, S.C.; Gayathri, S.R.; Chakravarthy, V.S. The Influence of Neural Activity and Neural Cytoarchitecture on Cerebrovascular Arborization: A Computational Model. Front. Neurosci. 2022, 16, 917196. [Google Scholar] [CrossRef] [PubMed]
- Lawn, T.; Howard, M.A.; Turkheimer, F.; Misic, B.; Deco, G.; Martins, D.; Dipasquale, O. From neurotransmitters to networks: Transcending organisational hierarchies with molecular-informed functional imaging. Neurosci. Biobehav. Rev. 2023, 150, 105193. [Google Scholar] [CrossRef] [PubMed]
- Lea-Carnall, C.A.; El-Deredy, W.; Stagg, C.J.; Williams, S.R.; Trujillo-Barreto, N.J. A mean-field model of glutamate and GABA synaptic dynamics for functional MRS. NeuroImage 2022, 266, 119813–119813. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Kim, Y.-K. Increased plasma VEGF levels in major depressive or manic episodes in patients with mood disorders. J. Affect. Disord. 2012, 136, 181–184. [Google Scholar] [CrossRef]
- Libby, J.; Marghoub, A.; Johnson, D.; Khonsari, R.H.; Fagan, M.J.; Moazen, M. Modelling human skull growth: a validated computational model. J. R. Soc. Interface 2017, 14, 20170202. [Google Scholar] [CrossRef]
- Lim, L.; Mi, D.; Llorca, A.; Marín, O. Development and functional diversification of cortical interneurons. Neuron 2018, 100, 294–313. [Google Scholar] [CrossRef]
- Lindenmayer, A. Mathematical models for cellular interactions in development. I. Filaments with one-sided inputs. J Theor Biol 1968, 18, 280–99. [Google Scholar]
- Litwin-Kumar, A.; Doiron, B. Formation and maintenance of neuronal assemblies through synaptic plasticity. Nat. Commun. 2014, 5, 5319–5319. [Google Scholar] [CrossRef]
- Liu, Q.-S.; Xu, Q.; Arcuino, G.; Kang, J.; Nedergaard, M. Astrocyte-mediated activation of neuronal kainate receptors. Proc. Natl. Acad. Sci. 2004, 101, 3172–3177. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, M.; Zhou, Y.; He, Y.; Hao, Y.; Song, M.; Yu, C.; Liu, H.; Liu, Z.; Jiang, T. Disrupted small-world networks in schizophrenia. Brain 2008, 131, 945–961. [Google Scholar] [CrossRef]
- Liu, Y.; Ouyang, P.; Zheng, Y.; Mi, L.; Zhao, J.; Ning, Y.; Guo, W. A Selective Review of the Excitatory-Inhibitory Imbalance in Schizophrenia: Underlying Biology, Genetics, Microcircuits, and Symptoms. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Lodato, S.; Rouaux, C.; Quast, K.B.; Jantrachotechatchawan, C.; Studer, M.; Hensch, T.K.; Arlotta, P. Excitatory Projection Neuron Subtypes Control the Distribution of Local Inhibitory Interneurons in the Cerebral Cortex. Neuron 2011, 69, 763–779. [Google Scholar] [CrossRef] [PubMed]
- Lord, L.-D.; Expert, P.; Huckins, J.F.; E Turkheimer, F.E. Cerebral Energy Metabolism and the Brain’s Functional Network Architecture: An Integrative Review. Br. J. Pharmacol. 2013, 33, 1347–1354. [Google Scholar] [CrossRef]
- Lord, L.-D.; Stevner, A.B.; Deco, G.; Kringelbach, M.L. Understanding principles of integration and segregation using whole-brain computational connectomics: implications for neuropsychiatric disorders. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2017, 375, 20160283. [Google Scholar] [CrossRef]
- Luppi, A.I.; Rosas, F.E.; Mediano, P.A.; Menon, D.K.; Stamatakis, E.A. Information decomposition and the informational architecture of the brain. Trends Cogn. Sci. 2024, 28, 352–368. [Google Scholar] [CrossRef]
- Mapelli, J.; Gandolfi, D.; Vilella, A.; Zoli, M.; Bigiani, A. Heterosynaptic GABAergic plasticity bidirectionally driven by the activity of pre- and postsynaptic NMDA receptors. Proc. Natl. Acad. Sci. 2016, 113, 9898–9903. [Google Scholar] [CrossRef]
- Marcaggi, P.; Attwell, D. Role of glial amino acid transporters in synaptic transmission and brain energetics. Glia 2004, 47, 217–225. [Google Scholar] [CrossRef]
- Markello, R.D.; Hansen, J.Y.; Liu, Z.-Q.; Bazinet, V.; Shafiei, G.; Suárez, L.E.; Blostein, N.; Seidlitz, J.; Baillet, S.; Satterthwaite, T.D.; et al. neuromaps: structural and functional interpretation of brain maps. Nat. Methods 2022, 19, 1472–1479. [Google Scholar] [CrossRef]
- Meier, M.H.; Shalev, I.; Moffitt, T.E.; Kapur, S.; Keefe, R.S.; Wong, T.Y.; Belsky, D.W.; Harrington, H.; Hogan, S.; Houts, R.; et al. Microvascular Abnormality in Schizophrenia as Shown by Retinal Imaging. Am. J. Psychiatry 2013, 170, 1451–1459. [Google Scholar] [CrossRef]
- Menassa, D.A.; Gomez-Nicola, D. Microglial Dynamics During Human Brain Development. Front. Immunol. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Meyer, H.S.; Schwarz, D.; Wimmer, V.C.; Schmitt, A.C.; Kerr, J.N.D.; Sakmann, B.; Helmstaedter, M. Inhibitory interneurons in a cortical column form hot zones of inhibition in layers 2 and 5A. Proc. Natl. Acad. Sci. 2011, 108, 16807–16812. [Google Scholar] [CrossRef] [PubMed]
- Milošević, N.T.; Ristanović, D. Fractality of dendritic arborization of spinal cord neurons. Neurosci. Lett. 2006, 396, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Min, R.; Nevian, T. Astrocyte signaling controls spike timing–dependent depression at neocortical synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, G.; Hjerling-Leffler, J.; Karayannis, T.; Sousa, V.H.; Butt, S.J.B.; Battiste, J.; Johnson, J.E.; Machold, R.P.; Fishell, G. Genetic Fate Mapping Reveals That the Caudal Ganglionic Eminence Produces a Large and Diverse Population of Superficial Cortical Interneurons. J. Neurosci. 2010, 30, 1582–1594. [Google Scholar] [CrossRef]
- Morigiwa, K.; Tauchi, M.; Fukuda, Y. Fractal analysis of ganglion cell dendritic branching patterns of the rat and cat retinae. Neurosci. Res. Suppl. 1989, 10, S131–S139. [Google Scholar] [CrossRef]
- Muldoon, S.F.; Bridgeford, E.W.; Bassett, D.S. Small-World Propensity and Weighted Brain Networks. Sci. Rep. 2016, 6, 22057. [Google Scholar] [CrossRef]
- Navarrete, M.; Perea, G.; de Sevilla, D.F.; Gómez-Gonzalo, M.; Núñez, A.; Martín, E.D.; Araque, A. Astrocytes Mediate In Vivo Cholinergic-Induced Synaptic Plasticity. PLOS Biol. 2012, 10, e1001259. [Google Scholar] [CrossRef]
- Nunes, F.D.D.; Ferezin, L.P.; Pereira, S.C.; Figaro-Drumond, F.V.; Pinheiro, L.C.; Menezes, I.C.; Baes, C.v.W.; Coeli-Lacchini, F.B.; Tanus-Santos, J.E.; Juruena, M.F.; et al. The Association of Biochemical and Genetic Biomarkers in VEGF Pathway with Depression. Pharmaceutics 2022, 14, 2757. [Google Scholar] [CrossRef]
- Ofer, N.; Shefi, O.; Yaari, G. Branching morphology determines signal propagation dynamics in neurons. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Okun, M.; Lampl, I. Instantaneous correlation of excitation and inhibition during ongoing and sensory-evoked activities. Nat. Neurosci. 2008, 11, 535–537. [Google Scholar] [CrossRef]
- Oldham, S.; Fulcher, B.D.; Aquino, K.; Arnatkevičiūtė, A.; Paquola, C.; Shishegar, R.; Fornito, A. Modeling spatial, developmental, physiological, and topological constraints on human brain connectivity. Sci. Adv. 2022, 8, eabm6127. [Google Scholar] [CrossRef] [PubMed]
- Öz, G.; Seaquist, E.R.; Kumar, A.; Criego, A.B.; Benedict, L.E.; Rao, J.P.; Henry, P.-G.; Van De Moortele, P.-F.; Gruetter, R.; Heinonen, I.; et al. Human brain glycogen content and metabolism: implications on its role in brain energy metabolism. Am. J. Physiol. Metab. 2007, 292, E946–E951. [Google Scholar] [CrossRef]
- Palmer, T.D.; Willhoite, A.R.; Gage, F.H. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol 2000, 425, 479–94. [Google Scholar] [CrossRef] [PubMed]
- Pandit, A.S.; Expert, P.; Lambiotte, R.; Bonnelle, V.; Leech, R.; Turkheimer, F.E.; Sharp, D.J. Traumatic brain injury impairs small-world topology. Neurology 2013, 80, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Papo, D.; Buldú, J. Does the brain behave like a (complex) network? I. Dynamics. Phys. Life Rev. 2023, 48, 47–98. [Google Scholar] [CrossRef]
- Páscoa dos Santos F, Verschure PF. 2024. Metastable Dynamics Emerge from Local Excitatory-Inhibitory Homeostasis in the Cortex at Rest. bioRxiv:2024.08.24.609501.
- Peyrounette, M.; Davit, Y.; Quintard, M.; Lorthois, S. Multiscale modelling of blood flow in cerebral microcirculation: Details at capillary scale control accuracy at the level of the cortex. PLOS ONE 2018, 13, e0189474. [Google Scholar] [CrossRef]
- Pla, R.; Stanco, A.; Howard, M.A.; Rubin, A.N.; Vogt, D.; Mortimer, N. Dlx1 and Dlx2 Promote Interneuron GABA Synthesis, Synaptogenesis, and Dendritogenesis. Cereb Cortex 2018, 28, 3797–3815. [Google Scholar] [CrossRef]
- Podvalny, E.; Noy, N.; Harel, M.; Bickel, S.; Chechik, G.; Schroeder, C.E.; Mehta, A.D.; Tsodyks, M.; Malach, R. A unifying principle underlying the extracellular field potential spectral responses in the human cortex. J. Neurophysiol. 2015, 114, 505–519. [Google Scholar] [CrossRef]
- Poil, S.S.; Hardstone, R.; Mansvelder, H.D.; Linkenkaer-Hansen, K. Critical-state dynamics of avalanches and oscillations jointly emerge from balanced excitation/inhibition in neuronal networks. J Neurosci 2012, 32, 9817–23. [Google Scholar] [CrossRef]
- Pope, M.; Fukushima, M.; Betzel, R.F.; Sporns, O. Modular origins of high-amplitude cofluctuations in fine-scale functional connectivity dynamics. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Pulvermüller, F.; Tomasello, R.; Henningsen-Schomers, M.R.; Wennekers, T. Biological constraints on neural network models of cognitive function. Nat. Rev. Neurosci. 2021, 22, 488–502. [Google Scholar] [CrossRef] [PubMed]
- Rae, C.D. A Guide to the Metabolic Pathways and Function of Metabolites Observed in Human Brain 1H Magnetic Resonance Spectra. Neurochem. Res. 2013, 39, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Ramezanian-Panahi, M.; Abrevaya, G.; Gagnon-Audet, J.-C.; Voleti, V.; Rish, I.; Dumas, G. Generative Models of Brain Dynamics. Front. Artif. Intell. 2022, 5, 807406. [Google Scholar] [CrossRef]
- Ramón y Cajal, S. Textura del sistema nervioso del hombre y de los vertebrados : estudios sobre el plan estructural y composición histológica de los centros nerviosos adicionados de consideraciones fisiológicas fundadas en los nuevos descubrimientos: Madrid : N. Moya, 1899; 1899-1904.
- Roca, C.; Adams, R.H. Regulation of vascular morphogenesis by Notch signaling. Genes Dev. 2007, 21, 2511–2524. [Google Scholar] [CrossRef]
- Rothman, D.L.; Behar, K.L.; Hyder, F.; Shulman, R.G. In vivo NMR Studies of the Glutamate Neurotransmitter Flux and Neuroenergetics: Implications for Brain Function. Annu. Rev. Physiol. 2003, 65, 401–427. [Google Scholar] [CrossRef]
- Rowitch, D.H.; Kriegstein, A.R. Developmental genetics of vertebrate glial–cell specification. Nature 2010, 468, 214–222. [Google Scholar] [CrossRef]
- Rubinov, M. Distinctions between brain structure, complexity, and function. Comment on “Does the brain behave like a (complex) network? I. Dynamics” by Papo and Buldú. Physics of Life Reviews 2024, 50, 30–31. [Google Scholar]
- Sahara, S.; Yanagawa, Y.; O'Leary, D.D.M.; Stevens, C.F. The Fraction of Cortical GABAergic Neurons Is Constant from Near the Start of Cortical Neurogenesis to Adulthood. J. Neurosci. 2012, 32, 4755–4761. [Google Scholar] [CrossRef]
- Santello, M.; Bezzi, P. ; VolterraA. TNFalpha controls glutamatergic gliotransmission in the hippocampal dentate gyrus. Neuron 2011, 69, 988–1001. [Google Scholar]
- Santello, M.; Cali, C.; Bezzi, P. Gliotransmission and the tripartite synapse. Adv Exp Med Biol 2012, 970, 307–31. [Google Scholar]
- Shanahan, M. Metastable chimera states in community-structured oscillator networks. Chaos: Interdiscip. J. Nonlinear Sci. 2010, 20, 013108–013108. [Google Scholar] [CrossRef] [PubMed]
- Shipley, R.J.; Smith, A.F.; Sweeney, P.W.; Pries, A.R.; Secomb, T.W. A hybrid discrete–continuum approach for modelling microcirculatory blood flow. Math. Med. Biol. A J. IMA 2019, 37, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Shonkoff JP, Phillips DA, National Research Council (U.S.). Committee on Integrating the Science of Early Childhood Development. 2000. From neurons to neighborhoods : the science of early child development. Washington, D.C.: National Academy Press. xviii, 588 p. p.
- Shu, Y.; Hasenstaub, A.; McCormick, D.A. Turning on and off recurrent balanced cortical activity. Nature 2003, 423, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.L.; Trachtenberg, J.T. Experience-dependent binocular competition in the visual cortex begins at eye opening. Nat. Neurosci. 2007, 10, 370–375. [Google Scholar] [CrossRef]
- Smith, T.G., Jr.; Lange, G.D.; Marks, W.B. Fractal methods and results in cellular morphology--dimensions, lacunarity and multifractals. J Neurosci Methods 1996, 69, 123–36. [Google Scholar] [CrossRef]
- Southwell, D.G.; Paredes, M.F.; Galvao, R.P.; Jones, D.L.; Froemke, R.C.; Sebe, J.Y.; Alfaro-Cervello, C.; Tang, Y.; Garcia-Verdugo, J.M.; Rubenstein, J.L.; et al. Intrinsically determined cell death of developing cortical interneurons. Nature 2012, 491, 109–113. [Google Scholar] [CrossRef]
- Sporns, O. The complex brain: connectivity, dynamics, information. Trends Cogn Sci 2022, 26, 1066–1067. [Google Scholar] [CrossRef]
- Sporns, O.; Kötter, R. Motifs in Brain Networks. PLOS Biol. 2004, 2, e369–e369. [Google Scholar] [CrossRef]
- Stam, C.J.; Jones, B.F.; Nolte, G.; Breakspear, M.; Scheltens, P. Small-world networks and functional connectivity in Alzheimer's disease. Cereb Cortex 2007, 17, 92–9. [Google Scholar] [CrossRef]
- Strogatz, S.H. Exploring complex networks. Nature 2001, 410, 268–276. [Google Scholar] [CrossRef]
- Takeda, T.; Ishikawa, A.; Ohtomo, K.; Kobayashi, Y.; Matsuoka, T. Fractal dimension of dendritic tree of cerebellar Purkinje cell during onto- and phylogenetic development. Neurosci. Res. 1992, 13, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Edelman, G.M.; Sporns, O. Complexity and coherency: integrating information in the brain. Trends in Cognitive Sciences 1998, 2, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Touboul, J.; Wendling, F.; Chauvel, P.; Faugeras, O. Neural mass activity, bifurcations, and epilepsy. Neural Comput 2011, 23, 3232–86. [Google Scholar] [CrossRef]
- Turkheimer, F.E.; Leech, R.; Expert, P.; Lord, L.-D.; Vernon, A.C. The brain's code and its canonical computational motifs. From sensory cortex to the default mode network: A multi-scale model of brain function in health and disease. Neurosci. Biobehav. Rev. 2015, 55, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Turkheimer, F.E.; Rosas, F.E.; Dipasquale, O.; Martins, D.; Fagerholm, E.D.; Expert, P.; Váša, F.; Lord, L.-D.; Leech, R. A Complex Systems Perspective on Neuroimaging Studies of Behavior and Its Disorders. Neurosci. 2021, 28, 382–399. [Google Scholar] [CrossRef]
- van Vreeswijk, C.; Sompolinsky, H. Chaos in Neuronal Networks with Balanced Excitatory and Inhibitory Activity. Science 1996, 274, 1724–1726. [Google Scholar] [CrossRef]
- van Vreeswijk, C.; Sompolinsky, H. Chaotic Balanced State in a Model of Cortical Circuits. Neural Comput. 1998, 10, 1321–1371. [Google Scholar] [CrossRef]
- Váša, F.; Shanahan, M.; Hellyer, P.J.; Scott, G.; Cabral, J.; Leech, R. Effects of lesions on synchrony and metastability in cortical networks. NeuroImage 2015, 118, 456–467. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Astroglial cradle in the life of the synapse. Philos Trans R Soc Lond B Biol Sci 2014, 369, 20130595. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol Rev 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Vértes, P.E.; Alexander-Bloch, A.F.; Gogtay, N.; Giedd, J.N.; Rapoport, J.L.; Bullmore, E.T. Simple models of human brain functional networks. Proc. Natl. Acad. Sci. 2012, 109, 5868–5873. [Google Scholar] [CrossRef] [PubMed]
- Vogels, T.P.; Sprekeler, H.; Zenke, F.; Clopath, C.; Gerstner, W. Inhibitory Plasticity Balances Excitation and Inhibition in Sensory Pathways and Memory Networks. Science 2011, 334, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-J.; Buzsáki, G. Gamma Oscillation by Synaptic Inhibition in a Hippocampal Interneuronal Network Model. J. Neurosci. 1996, 16, 6402–6413. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, S.; Li, M. Neurovascular crosstalk and cerebrovascular alterations: an underestimated therapeutic target in autism spectrum disorders. Front. Cell. Neurosci. 2023, 17, 1226580. [Google Scholar] [CrossRef]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Schautz, B.; Later, W.; Heymsfield, S.B.; Müller, M.J. Specific metabolic rates of major organs and tissues across adulthood: evaluation by mechanistic model of resting energy expenditure. Am. J. Clin. Nutr. 2010, 92, 1369–1377. [Google Scholar] [CrossRef]
- Waschke, L.; Donoghue, T.; Fiedler, L.; Smith, S.; Garrett, D.D.; Voytek, B.; Obleser, J. Modality-specific tracking of attention and sensory statistics in the human electrophysiological spectral exponent. eLife 2021, 10. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Wearne, S.; Rodriguez, A.; Ehlenberger, D.; Rocher, A.; Henderson, S.; Hof, P. New techniques for imaging, digitization and analysis of three-dimensional neural morphology on multiple scales. 2005, 136, 661–680. [Google Scholar] [CrossRef]
- Wigström, H.; Gustafsson, B. Facilitation of hippocampal long-lasting potentiation by GABA antagonists. Acta Physiol. Scand. 1985, 125, 159–172. [Google Scholar] [CrossRef]
- Wu, G.K.; Tao, H.W.; Zhang, L.I. From elementary synaptic circuits to information processing in primary auditory cortex. Neurosci. Biobehav. Rev. 2011, 35, 2094–2104. [Google Scholar] [CrossRef]
- Wu, M.W.; Kourdougli, N.; Portera-Cailliau, C. Network state transitions during cortical development. Nat. Rev. Neurosci. 2024, 25, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Stathopoulou, M.G.; de Andrés, F.; Siest, G.; Murray, H.; Martin, M.; Cobaleda, J.; Delgado, A.; Lamont, J.; Peñas-Liedó, E.; et al. VEGF-related polymorphisms identified by GWAS and risk for major depression. Transl. Psychiatry 2017, 7, e1055. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.C.; Vest, R.T.; Kern, F.; Lee, D.P.; Agam, M.; Maat, C.A.; Losada, P.M.; Chen, M.B.; Schaum, N.; Khoury, N.; et al. A human brain vascular atlas reveals diverse mediators of Alzheimer’s risk. Nature 2022, 603, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; La Camera, G. Co-existence of synaptic plasticity and metastable dynamics in a spiking model of cortical circuits. PLoS Comput Biol 2024, 20, e1012220. [Google Scholar]
- Yang, Y.; Guan, W.; Sheng, X.-M.; Gu, H.-J. Role of Semaphorin 3A in common psychiatric illnesses such as schizophrenia, depression, and anxiety. Biochem. Pharmacol. 2024, 226, 116358. [Google Scholar] [CrossRef]
- Yarkoni, T.; A Poldrack, R.; E Nichols, T.; Van Essen, D.C.; Wager, T.D. Large-scale automated synthesis of human functional neuroimaging data. Nat. Methods 2011, 8, 665–670. [Google Scholar] [CrossRef]
- Ye, Z.; Mostajo-Radji, M.A.; Brown, J.R.; Rouaux, C.; Tomassy, G.S.; Hensch, T.K. Instructing Perisomatic Inhibition by Direct Lineage Reprogramming of Neocortical Projection Neurons. Neuron 2015, 88, 475–83. [Google Scholar]
- Zang, Y.; Chaudhari, K.; Bashaw, G.J. New insights into the molecular mechanisms of axon guidance receptor regulation and signaling. Curr Top Dev Biol 2021, 142, 147–196. [Google Scholar]
- Zhang S, Larsen B, Sydnor VJ, Zeng T, An L, Yan X and others. 2023. In-vivo whole-cortex estimation of excitation-inhibition ratio indexes cortical maturation and cognitive ability in youth. bioRxiv:2023.06.22.546023.
Figure 1.
Alternative architectures of elementary feedback systems that enable the generation of fast frequency waves, the building blocks of brain functional connectivity. Excitatory units are constituted by pyramidal cells while inhibitory cells are GABA interneurons.
Figure 1.
Alternative architectures of elementary feedback systems that enable the generation of fast frequency waves, the building blocks of brain functional connectivity. Excitatory units are constituted by pyramidal cells while inhibitory cells are GABA interneurons.
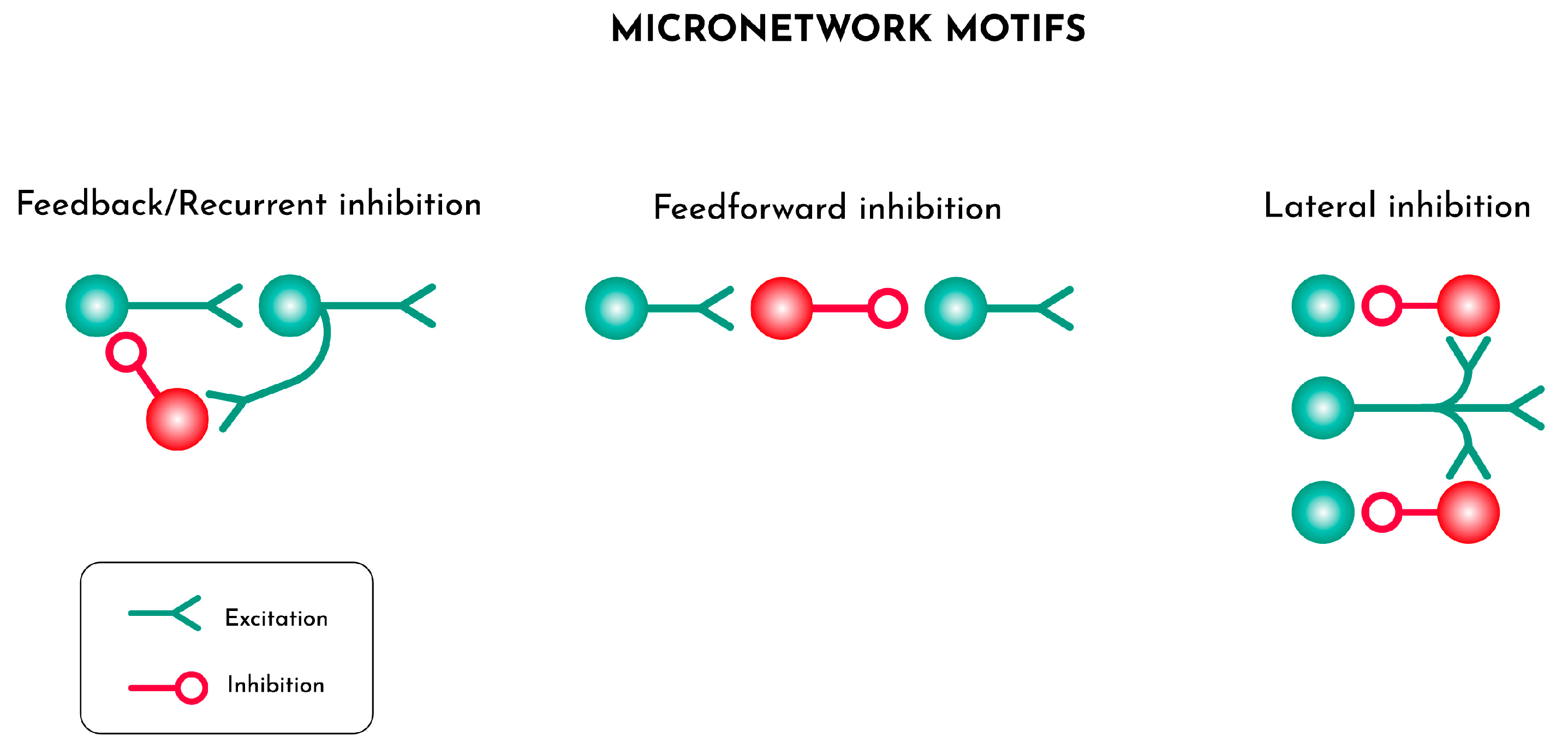
Figure 2.
From left to right, the various stages of cortical formation during embryogenesis up to the postnatal brain. Neural stem/progenitor cells (NSCs/NPCs) start a first differentiation stage mostly converting into radial glial cells (RGCs) that make the scaffolding starting from the ventricular up to the pial surface while novel blood vessels grow inwards. Local metabolic, ionic and molecular conditions then drive the generation and migration of glial and neuronal cells and their connections to the developing vascular networks. Finally, the same local conditions promote the differentiation and maturation of oligodendrocytes (OL) from their progenitor cells (OLP) and axonal myelination.
Figure 2.
From left to right, the various stages of cortical formation during embryogenesis up to the postnatal brain. Neural stem/progenitor cells (NSCs/NPCs) start a first differentiation stage mostly converting into radial glial cells (RGCs) that make the scaffolding starting from the ventricular up to the pial surface while novel blood vessels grow inwards. Local metabolic, ionic and molecular conditions then drive the generation and migration of glial and neuronal cells and their connections to the developing vascular networks. Finally, the same local conditions promote the differentiation and maturation of oligodendrocytes (OL) from their progenitor cells (OLP) and axonal myelination.
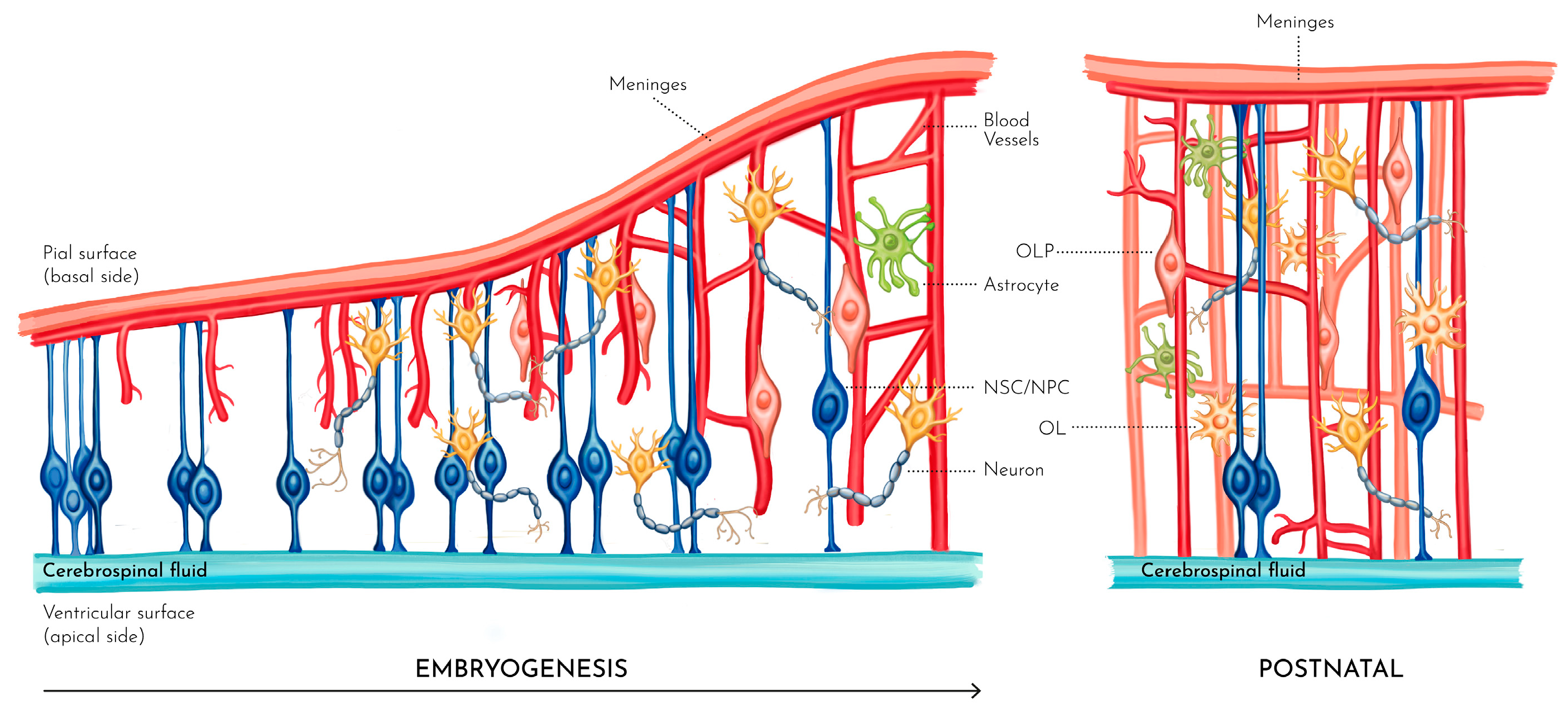
Figure 3.
The neurogliovascular unit is here presented as the elementary unit for the better modelling of brain morphology and function using generative computations.
Figure 3.
The neurogliovascular unit is here presented as the elementary unit for the better modelling of brain morphology and function using generative computations.
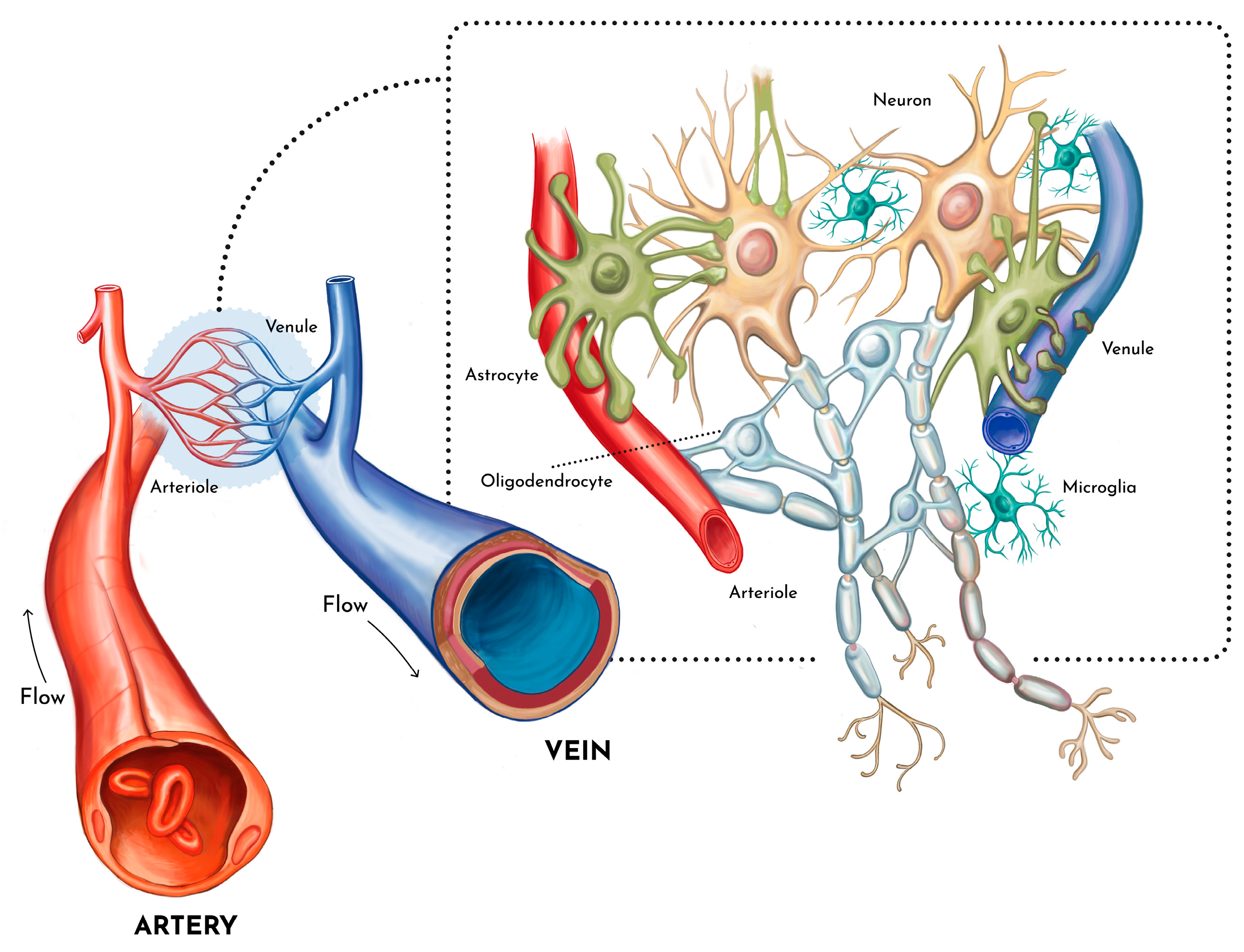
Figure 4.
The process of vascular branching in brain follows to feedback systems. A) Astrocytes (round cells) sense hypoxic conditions caused by expanding neural networks (blue) and release VEGF that stimulates sprouting of endothelial cells from the vascular networks (red). B) Strongest emerging tubes from endothelial cells inhibit further sprouting from neighbouring cells.
Figure 4.
The process of vascular branching in brain follows to feedback systems. A) Astrocytes (round cells) sense hypoxic conditions caused by expanding neural networks (blue) and release VEGF that stimulates sprouting of endothelial cells from the vascular networks (red). B) Strongest emerging tubes from endothelial cells inhibit further sprouting from neighbouring cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



