
Submitted:
17 December 2024
Posted:
18 December 2024
You are already at the latest version
Abstract
Classical Rett syndrome (RTT) is a neurodevelopmental disorder caused by mutations in the MECP2 gene, resulting in a devastating phenotype associated with a lack of gene expression control. Mouse models lacking Mecp2 expression with an RTT-like phenotype have been developed to advance therapeutic alternatives. Environmental enrichment (EE) attenuates RTT symptoms in patients and mouse models. However, the mechanisms underlying the effects of EE on RTT have not been fully elucidated. We housed male hemizygous Mecp2-null (Mecp2-/y) and wild-type mice in specially conditioned cages to enhance sensory, cognitive, social, and motor stimulation. EE attenuated the progression of the RTT phenotype by preserving neuronal cytoarchitecture and neural plasticity markers. Furthermore, EE ameliorated defects in neuromuscular junction organization and restored the motor deficit of Mecp2-/y mice. Treatment with plasma from young WT mice was used to assess whether the increased activity could modify plasma components, mimicking the benefits of EE in Mecp2-/y. Plasma treatment attenuated the RTT phenotype by improving neurological markers, suggesting that peripheral signals of mice with normal motor function have the potential to reactivate dormant neurodevelopment in RTT mice. These findings demonstrate how EE and treatment with young plasma ameliorate RTT-like phenotype in mice, opening new therapeutical approaches for RTT patients.
Keywords:
Rett syndrome
; Mecp2
; Environmental enrichment
; Plasma treatment
1. Introduction
Classical Rett syndrome (RTT, OMIM #312750) is a devastating neurodevelopmental disorder and one of the leading causes of cognitive deficit in young women, with an incidence of 1 in 5,000-10,000 female births [1,2]. RTT patients have a seemingly normal neurological development during the first 6-18 months of life, followed by stagnation, and later fall into a developmental regression accompanied by the onset of symptoms such as motor impairment, loss of hand skills, seizure, autonomic dysfunction, anxiety alterations, and intellectual disabilities, among other neurological manifestations [3,4,5]. In addition to these nervous system-associated phenotypes, RTT patients also exhibit peripheral alterations such as respiratory abnormalities [6], evidence of axonopathy with skeletal muscle alterations as a probable consequence of denervation [7]. Cardiorespiratory failure has been reported as one of the most prevalent causes of death in RTT patients [8].
Mutations in the X-linked gene MECP2 are the main cause of RTT [3,9,10]. MECP2 encodes for two isoforms of the Methyl CpG Binding Protein-2 (MECP2), a transcriptional regulator that binds to methylated and hydroxymethylated cytosine in CpG dinucleotides to recruit transcriptional regulatory complexes to either decrease or increase the expression of its target genes [11,12]. At the time of birth, MECP2 expression is low. Still, its expression gradually increases until it reaches the highest levels in mature neurons [13,14], which explains the RTT phenotype as neurodevelopment progresses and highlights the role of MECP2 as a DNA methylation/hydroxymethylation reader critical to maintaining neuronal functions in the mature brain.
Due to the monogenetic nature of Rett syndrome, several mouse models lacking Mecp2 expression have been developed to investigate the pathophysiological and cellular bases of RTT and uncover the molecular mechanisms underlying the RTT phenotype [15,16,17]. These mouse models recapitulate most of the RTT phenotype observed in RTT patients and have been extensively characterized [15,16,18,19]. Studies using RTT mouse models have shown that neurons lacking the expression of Mecp2 postnatally exhibit altered maintenance of the mature neuronal network [20]. In addition, conditional re-expression of Mecp2 in adult hemizygous Mecp2-null mutant (Mecp2-/y) mice rescues most RTT phenotypes [21,22,23,24]. This evidence demonstrates that Mecp2 is required to maintain neuronal function and that its absence does not irreversibly affect the arrested neurodevelopment observed in RTT. This evidence provides an opportunity for developing potential therapeutic interventions for RTT patients.
Evidence shows that RTT-associated phenotype may result from an alteration in dendritic complexity and spine dysgenesis [25]. Indeed, postmortem fixed brains from RTT patients exhibit reduced dendritic complexity and decreased spine density [3,26,27,28]. Moreover, iPSC-derived neurons from RTT patients exhibit defects in neuronal maturation and synaptic formation [29,30]. As noted, RTT-mouse models offer an excellent platform for studying brain alterations associated with RTT, as they replicate most of the phenotypes in RTT patients. For instance, the decreased synaptic parameters observed in RTT patients have also been observed in layer II/III of the motor cortex and layer V of the somatosensory cortex of Mecp2-/y mice [31,32,33,34]. In addition, the number of excitatory synapses observed in primary cortical cultures of Mecp2-knockdown mice is reduced compared to wild-type mice [35]. These findings support the hypothesis that impaired synaptic maturation underlies the neuronal dysfunction observed in RTT. Thus, interventions to prevent or reverse neural decline may represent promising therapeutic approaches for RTT patients.
Environmental enrichment (EE) is an experimental paradigm widely used in mouse models to enhance neuronal plasticity and modulate the pathogenesis of central nervous system disorders through molecular, cellular, and behavioral effects [36]. EE has shown promising results in RTT mouse models and has even been proposed as a therapeutic alternative not only for RTT patients [37], but also for other neurodevelopmental disorders [37]. This hypothesis is supported by evidence from independent groups using different RTT mouse models. Kondo et al. demonstrated that EE ameliorates the motor coordination deficits of heterozygous Mecp2-null females exposed to EE for 4 weeks; however, this paradigm was ineffective in Mecp2-/y males [38]. Subsequently, it was demonstrated that EE attenuates locomotor deficit in Mecp2-/y male mice exposed to EE since weaning, likely by increasing brain matter [39] and by inducing an unconventional transcriptional response that is not associated with enhanced expression of synaptic markers in mice exposed temporarily to an EE housing [40]. These studies underscore the importance of an early intervention to attenuate the progression of RTT-like symptoms. Lonetti et al. demonstrated that EE promotes synaptic plasticity and synapse formation in Mecp2-/y male mice exposed to EE from 10 days of age before RTT-related phenotypes are evident [41]. Mecp2-/y mice exhibit decreased locomotor activity and neuronal plasticity [40,42], and EE includes elements that encourage increased locomotor activity, which has been described to enhance circulating levels of neurotrophins, potentially impacting the central nervous system [43,44]. Moreover, treatment with young plasma effectively rescues the attenuated neural plasticity observed in aged mice [45]. These findings suggest that elements in the plasma of mice with regular motor activity might reflect changes in the plasma of Mecp2-/y mice exposed to EE.
We aimed to gain insight into the mechanism by which EE promotes brain gain of function and attenuates the RTT-like phenotype in a mouse model of the disease with the goal of designing plausible intervention strategies for patients. To this end, we housed Mecp2-/y and wild-type (WT) male mice in specially conditioned cages to enhance sensory, cognitive, social, and motor stimulation. We compared behavioral, molecular, and cellular parameters between Mecp2-/y mice exposed to EE and those housed in regular conditions. We found that continuous exposure to EE attenuates the progression of the RTT phenotype by preventing damage to neuronal cytoarchitecture. These effects were partially replicated by intraperitoneal plasma injection from healthy young mice. These results show that early and permanent exposure to EE attenuates the RTT-phenotype progression by a mechanism associated with preventing cytoarchitecture deterioration. Besides, this EE-induced effect was partially emulated by treatment with plasma from WT young mice, indicating that peripheral signals present in mice with regular motor activity prevent brain cytoarchitecture deterioration and attenuate the RTT progression.
2. Materials and Methods
2.1. Mice, Housing Conditions, and Genotyping
To determine the effect of EE exposure on RTT-like phenotype, we used the Mecp2-/y mouse line generated by Adrian Bird’s lab [15]. Considering the fertility and maternal care provided by females in a 129/SvJ genetic background, heterozygous Mecp2-null females in a genetic background C57BL/6 obtained from Jackson Laboratory stock #003890 were bred with 129/SvJ wild-type males for at least 10 generations. Then, we generate Mecp2-/y male mice in a mixed homogeneous C57BL/6:129SvJ genetic background obtained by mating heterozygous Mecp2-null 129SvJ females with WT C57BL/6 males.
The genotype of mice was determined by PCR analysis of DNA extracted from a tail biopsy of 14-21 days of age to identify transgenic mice using the following primers: F: CCACCCTCCAGTTTGGTTTA, R1: GACCCCTTGGGACTGAAGTT, and R2: CCATGCGATAAGCTTGATGA. At weaning, Mecp2-/y mice and their WT littermates were randomly housed in either standard conditions (SC) or environmental enrichment cages (EE) until being evaluated. SC comprises 3-4 mice housed in a 30x15 cm cage provided with bedding and ad libitum access to food (SD, Envigo S2019 and LabDiet 5P00 Prolab-RMH-3000) and water, whereas EE cages consisted of two connected 30x30 cm cages housing 7-8 mice with access to different bedding material, daily changed plastic toys, a free-running wheel, and ad libitum access to water and food contained in various containers and located in different places. Both types of cages were in the same room. All protocols were designed according to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication no. 85-23, revised 2011) and approved by the Centro de Estudios Científicos Animal Care and Use Committee.
2.2. Overall Phenotype and Behavioral Tests
To evaluate the RTT-like phenotype progression, mice in both SC or EE were assessed weekly in their overall state starting from 4 weeks of age by measuring body weight, lifespan, hindlimb clasping, tremor, and coat condition and according to that previously described [40,42,46]. Besides, the presence of ataxia in the gait as a consequence of cerebellar dysfunction in motor coordination was evaluated through a ledge test. The level of RTT-phenotype severity was determined according to an arbitrary scaling from 0 to 3, in which 0 means absence of the phenotype, and 3 a severe phenotype [42]. Clasping: 0, no clasping; 1, reversible clasping; 2, delayed but irreversible; 3, immediate and irreversible. Tremor: 0, no tremor; 1, slight and intermittent tremor; 2, permanent tremor; 3, moderate or severe tremor. Coat condition: 0, shiny and tidy coat; 1, partly oily or slightly messy coat; 2, oily and slightly messy coat; 3, oily and messy coat. Ledge test: 0, smooth edge displacement; 1, displacement with static periods; 2, displacement with difficulty and hindlimb slipped; 3, forelimb and hindlimb slipped from the edge. The scores obtained in each of the evaluations were summed to obtain a total score representing the general state of the mice and the RTT-phenotype progression.
At seven weeks of age, mice were daily evaluated for three consecutive days in a battery of behavioral tests, including plus maze, open field, hanging wire, and elevated dowel test to determine the RTT-like behavior progression as previously described [40,42] and briefly described as follow:
Plus Maze: To evaluate anxiety-like behavior and spatial perception, mice were placed in the center of an elevated maze with a cross of two open and two closed arms (homemade). The time spent in either the arms or the center of the maze was recorded. Open Field: To evaluate anxiety-like behavior, motor rearing, and exploratory activity, mice were placed in an open arena with a photo beam system (Med associated Inc). Mice were placed at the center of the arena, and their activity was recorded during 30 min at 10 min intervals. Hanging wire: To evaluate forepaw strength, the hanging wire test was performed by hanging mice by their forepaws from a suspended wire, and the number of falls in two minutes was recorded. Elevated dowel: To evaluate motor coordination, mice were assessed in a dowel apparatus consisting of two 70 cm elevated platforms enclosed by 7 cm high walls except for one side that connects both elevated platforms by a 70 cm long dowel of 0.7 cm radius. As described previously [40], mice were habituated by 1 min to each platform, then received a short training placing them on the dowel 10 cm away from one of the platforms. After habituation, mice were placed in the middle of the dowel, and the latency to start moving, time of the first arrival, the number of arrivals in a period of 90 sec, and the total number of falls recorded.
At eight weeks of age, mice were subjected to rotarod test during 4 consecutive days at nine weeks of age. Rotarod: To evaluated motor coordination’s and motor learning, mice were evaluated in accelerating rotarod for 4 consecutive days, 4 trials per day with intervals of 30 min of resting. The rotarod was configurated in accelerating mode, starting at 5 rpm to reach 20 rpm at 180 sec, and maintained for a maximum of 300 sec of total training. Mice were placed on the rotating cylinder, and the time to fall was recorded.
2.3. Golgi Staining and Morphological Evaluation
To determine the effect of the exposure to environmental enrichment on motor cortical cytoarchitecture, mice were deeply anesthetized and then transcardiac perfused with cold saline followed by 4% PFA. After that, the brain was removed from the skull and immersed in Golgi impregnation solution according to that described by the manufacturer of FD Rapid GolgiStainTMKit. The brains were sliced with a vibratome at 200 µm before the final staining and then stained sections were mounted with Vectamount in slides. Pyramidal neurons of layer V of motor cortex M1-M2 were drawn under a microscope, emulating the neurolucida system. To be included in the analysis, the neuronal soma should have a characteristic shape, be in the plane of the slide, and have the dendritic tree in the thick of the slide. Then, Sholl analysis was performed using an Image J plugin considering concentric circles of 20 µm. To determine the dendritic spine density, dendrites of 2nd and 3rd order were selected, and pictures were taken using an Olympus IX-71 microscope (Olympus, Germany) connected to an MSHOT digital camera. Images were analyzed with the software, and the dendritic density was evaluated with the ImageJ plugin Analyze Skeleton in a dendritic fragment of 30 µm.
2.4. RNA-Seq and Gene Ontology Enrichment Analysis
Brains from 7-week-old WT and Mecp2-/y mice were dissected. According to manufacturer’s instructions, RNA was isolated from forebrain with TRIzol (Invitrogen). A pool of four RNA samples per condition (approximately 4 mg of WT RNA and 4 ug of Mecp2-/y RNA) was sent to Macrogen Co., Ltd. as single replicate for quality control of total RNA integrity and sequencing by microarray using an Agilent Technologies 2100 Bioanalyzer (Agilent RNA 6,000 nano kit (Agilent, cat.# 5067-1511)). The basic statistics (Fold change, group mean, standard deviation), identification of differentially expressed genes (T-test, LPE test, ANOVA with p-value < 0.05), and multiple testing correction (Fold Discovery Rate, Bonferroni with adjusted p-value < 0.05) were carried out by the Analysis Service of Macrogen Co., Ltd.
Based on the differentially expressed genes, Gene Ontology Enrichment Analysis was performed using the ClueGO app in Cytoscape, using the following reference database: GO_ImmuneSystemProcess-EBI-UniProt-GOA-ACAP-ARAP_25.05.2022_00h00 : 3113. Default parameters were used.
2.5. Gene and Protein Expression
Gene expression: Brains were dissected from 7-week-old mice exposed to environmental enrichment or control cages for 5 weeks. RNA was isolated from the motor cortex and reverse transcribed as previously described [47]. Briefly, motor cortex was dissected, and samples were homogenized in TRIzol (Invitrogen) according to the manufacturer’s instructions. RNA was precipitated and treated with one unit of DNase I (Life Technologies). Five micrograms of total RNA were reverse transcribed using random primers and ImProm II kit (Promega). cDNA was quantified by qPCR using Kapa SYBR Quantimix (Kapa). The qPCR analysis was performed in triplicates from one reverse transcribed product using the Rotor-Gene 6000 (Corbett). Values were analyzed following the 2−ΔΔCt method using cyclophilin-A (Cyc) as a normalizer [48]. The list of primers that were used are described in Table 1. Protein expression: Brains were dissected, and the motor cortex was homogenized using a Douncer tissue grinder in RIPA buffer (Thermo Scientific) supplemented with 1x protease inhibitor cocktail (Sigma, P8340) and 1x phosphatase inhibitor cocktail (Pierce). Twenty-five micrograms of protein were electrophoresed on 4% and 8-12% SDS polyacrylamide gels, transferred onto nitrocellulose membranes (Bio-Rad), and blocked for 1 h at room temperature with freshly prepared TBS-T buffer containing 5% non-fat dry milk. Membranes were incubated overnight with anti-EEAT1 (GLAST) antibody (ab416, Abcam), anti-EEAT2 (GLT-1) antibody (sc-15317, Santa Cruz Biotechnology) or b-Actin antibody (sc-47778, Santa Cruz Biotecnology) at 4ºC, washed and incubated with secondary HRP-conjugated IgG for 2 h at room temperature. Bands were visualized with WESTAR SUPERNOVA Cod. XLS3 (Cyanagen, Italy) chemiluminescent substrate according to the manufacturer’s instructions in Syngene G:Box (UK). Densitometry of immunoreactive bands was quantitated with ImageJ software, using the expression of b-Actin as a normalizer.
2.6. Collection of Mouse Young Plasma Samples and Plasma Injection
6-week-old wild-type mice in genetic background C57BL/6 and 129/SvJ mice were deeply anesthetized by an intraperitoneal (IP) injection of Avertin (200 mg/Kg). Blood was extracted by cardiac function using a 22 G syringe, and blood was transferred to a 1,6 mL heparinized tube. Then, blood samples were centrifuged for 20 min at 2,000 x g, and the supernatant was centrifuged for 2 min at 14,000 x g to remove the platelet. After that, samples were freeze at -80ºC in 100 µL aliquots containing 50% C57BL/6 and 50% 129/SvJ mice plasma. 100 µL of young plasma was IP injected to Mecp2-/y mice, every other day from 4 weeks of age, with a total of 8 injections until 6 weeks of age.
2.7. Diaphragm Neuromuscular Junction
The diaphragm muscle was dissected, and the whole-mount was fixed at 0.5% formaldehyde (FA) in 1X Phosphate Buffered Saline (PBS) at 22 °C for 90 min. Samples were incubated with 0.1 M glycine in 1X PBS, permeabilized with PBST (1X PBS/0.5% TritonX-100), and blocked with 4% goat serum (GS) dissolved in PBST 1 h at 22 °C. Muscles were incubated with 4% GS-PBST containing Alexa488-conjugated α-bungarotoxin (BTX) (Invitrogen, Carlsbad, CA, USA) (1:500) 12-16 h at 4 °C. Samples were post-fixed with 0.5% FA in 1X PBS for 10 min at 22 °C and subsequently flat-mounted between two coverslips. For endplate band quantification, fluorescent images of a-BTX-stained right hemidiaphragm were captured with a ZEISS Axio Zoom.V16 scope, and AChR clusters distribution were analyzed. The endplate bandwidth of 20–40 bins per animal was measured with ImageJ.
2.8. Statistical Analysis
The software GraphPad PRISM Version 10.2.0 (San Diego, CA) was used for statistical analysis. Data are presented as mean ± SEM values, and differences were analyzed with Mann-Whitney test, one-way ANOVA, two-way ANOVA, or Simple survival analysis Mantel-Cox, as indicated in each figure. Statistical significances correspond to ✱p <0.05, ✱✱p <0.01, ✱✱✱p <0.001 and ✱✱✱✱p <0.0001, ns, non-statistical differences.
3. Results
3.1. The Exposure to a Neuronal Plasticity-Dependent Paradigm Ameliorates the Phenotype Exhibited by a Mouse Model of Rett Syndrome
RTT is characterized by a decreased neural plasticity and exposure to an experience-dependent paradigm has been shown to increase dendrite complexity in several mouse models of neurological disorders, among them RTT [40,49]. To gain insight into the cellular and molecular mechanism underlying this therapeutic effect, we permanently housed Mecp2-/y mice and their WT littermate in EE or SC since weaning, when the RTT phenotype if it is apparent is mild. We weekly recorded mouse weight and survival, and at 7 weeks of age, the general mouse condition was estimated by using a battery of phenotypic evaluation consisting of hindlimb clasping reflex and corporal tremor as indicators of neurological deterioration progression, piloerection as an indicator of grooming and social interaction, ledge test as a measurement of motor coordination, and the total score obtained from the total sum of each of the phenotypic evaluation was used as an indicator of mouse general condition.
Before puberty, Mecp2-/y mice showed decreased body weight, a phenotype that is inverted after puberty, as it is demonstrated by our results and evidence from the literature [4,15,16,50]. Mecp2-/y mice exposed to EE showed a reduced body weight compared to Mecp2-/y mice housed in control conditions (Figure 1A), indicating that the exposure to EE affects energy homeostasis. Furthermore, the decreased lifespan of Mecp2-/y mice was shown to be increased by exposure to EE. Mecp2-/y mice showed a half-life of 12 weeks of age, and no mice survived beyond 16 weeks; however, Mecp2-/y mice exposed to EE showed a half-life of 17 weeks (Figure 1B).
To evaluate the effect of EE exposure on RTT classic phenotype after puberty, mice were evaluated in a battery of general condition test. As expected, Mecp2-/y mice have an increased level of clasping compared to WT, but this neurological phenotype was not prevented by exposing mice to EE (Figure 1C). Despite this result, exposure to EE prevented the other phenotypic parameters evaluated. Mecp2-/y mice showed increased levels of corporal tremor, piloerection, and hindlimb discoordination compared to WT mice; however, Mecp2-/y mice exposed to EE showed a reduction in these phenotypes which was not different from WT mice, in contrast to Mecp2-/y mice in SC (Figure 1D, E, and F). As an evaluation of the general mice condition, the score of each test was summed to obtain a phenotypic evaluation total score, which was, as expected, higher in Mecp2-/y mice in comparison to WT mice; however, this parameter was rescued in Mecp2-/y mice exposed to EE compared to Mecp2-/y mice in SC. Together, these results show that permanent exposure to an EE since weaning improves energy homeostasis and increases lifespan at the time that attenuates the RTT-like phenotype.
3.2. The Neuronal Plasticity-Dependent Paradigm Exposure Reduces the Behavioral Alterations and Motor Deficits Exhibited by a Mouse Model of Rett Syndrome
The next step was to evaluate whether exposure to EE could attenuate the behavioral phenotype of the RTT mouse model. Mecp2-/y mice and their WT littermates were evaluated using a second battery of phenotypic evaluation to assess anxiety-like behavior, spatial perception, locomotion, and motor coordination performance. First, mice were assessed in the elevated plus maze. As expected, WT mice showed a marked preference for staying in the closed arms of the maze. As previously demonstrated [40], Mecp2-/y mice exposed to SC showed a higher preference for open arms and, on the contrary, less preference for closed arms, with no preference for any of the arms. EE exposure did not affect the arm preference in WT mice; however, in Mecp2-/y mice, EE reestablished the preference for closed arms (Figure 2A). Then, mice were evaluated in the open field test. We measured traveled distance as a locomotion parameter. Mecp2-/y mice exposed to SC showed significantly less locomotion activity than WT mice exposed to SC and EE. However, Mecp2-/y mice exposed to EE showed increased locomotor activity compared to Mecp2-/y mice exposed to SC (Figure 2B). Thus, EE exposure improved locomotion in an RTT mouse model.
A distinctive symptom of RTT patients is reduced muscle tone and motor control. Therefore, mice were evaluated in the wire-hanging test to assess whether EE exposure improves motor strenght. As expected, Mecp2-/y mice fail more often when hanging from an elevated wire than WT mice in SC. Mecp2-/y mice exposed to EE showed less failure when hanging from the wire than Mecp2-/y mice in SC, though higher than that observed in WT mice (Figure 2C). To determine the effect of EE exposure on motor coordination, function, and control, the performance of WT and RTT mice was evaluated in the elevated dowel test for 60 seconds. Mecp2-/y mice in SC showed a higher number of falls in comparison to WT mice, and this phenotype was entirely prevented by EE exposure since Mecp2-/y mice in EE showed a similar number of falls than WT mice and fewer falls than Mecp2-/y mice housed in SC (Figure 2D). The latency of mice to start moving and the time of the first arrival were also recorded. The results show that Mecp2-/y mice in SC started moving later than WT mice and took longer to reach the safe elevated platform. EE exposure improved these parameters in Mecp2-/y mice since the latency time and the time of the first arrival were similar to those exhibited by WT mice, and in the case of the first arrival, it was less than in Mecp2-/y mice in SC (Figure 2E and F). Following these results, the number of arrivals of Mecp2-/y mice in SC was less than in WT mice, and this phenotype was prevented entirely by EE exposure since Mecp2-/y mice in EE showed a similar number of arrivals as the WT mice and higher than Mecp2-/y mice in SC (Figure 2G). Motor coordination was also evaluated on the rotating cylinder of a rotarod, which is also used to assess motor learning. Mecp2-/y mice showed poor motor coordination and failure to stay on the rotating cylinder compared to WT mice. This motor phenotype was prevented entirely by exposure to EE since Mecp2-/y mice exposed to EE showed a motor phenotype comparable to WT mice in SC in the first training day. Despite this motor improvement, EE exposure failed to induce motor learning in Mecp2-/y mice (Figure 2H).
3.3. The Exposure to a Neuronal Plasticity-Dependent Paradigm Decreases the Synaptic Deficit Exhibited by an RTT Mouse Model
Since the general condition of the mice and behavioral tests showed that the EE exposure improved motor performance in Mecp2-/y mice, we next evaluated the motor cortex phenotype. To that, we compared the M1-M2 motor cortex cytoarchitecture at 3 and 7 weeks of age in WT and Mecp2-/y mice exposed to SC or EE housing. We chose these ages as critical times in RTT phenotype progression. While the RTT phenotype is barely apparent in mice at 3 weeks of age, at 7 weeks of age, motor performance is diminished in Mecp2-/y mice in SC, and in response to EE an improvement was observed at this age. At 3 weeks of age, the motor cortex of Mecp2-/y mice is not significantly different from WT mice (Figure 3A). However, at 7 weeks of age, the Mecp2-/y motor cortex seems disorganized, with reduced neuronal complexity and signs suggesting astrogliosis (Figure 3B). Nevertheless, the motor cortex of Mecp2-/y mice exposed to EE seems similar to WT mice (Figure 3C), with an apparent better organization, improved neuronal complexity, and no signs of astrogliosis, which contrasts with that observed in Mecp2-/y mice in SC (Figure 3B-C).
The dendritic length of cortical layer V pyramidal neurons was evaluated to compare this observation. The results show a reduction in the dendritic length of pyramidal neurons from Mecp2-/y mice in SC compared to those of WT mice. However, this difference was not observed in the dendritic length of Mecp2-/y mice exposed to EE (Figure 3D-E). The number of dendrites was also evaluated, and similar results were found. The pyramidal neurons of the motor cortex from Mecp2-/y mice in SC exhibit a decreased number of dendrites compared to those from WT mice. Nevertheless, the number of dendrites in pyramidal neurons of Mecp2-/y mice exposed to EE was similar to that of WT mice in SC (Figure 3F). These morphological changes related to neuronal plasticity were similar when the dendritic spine density was evaluated. Second-order dendrites of pyramidal neurons from the motor cortex of Mecp2-/y mice exhibit less spine density than those from WT mice in SC. However, in Mecp2-/y mice exposed to EE, the dendritic spine density was similar to that of WT mice in SC and higher than those of Mecp2-/y mice in SC (Figure 3G-H). These results show that early EE exposure increases structural neuronal plasticity markers and prevents the cortical impairment associated with RTT neurological progression.
3.4. The Expression of Genes Related to Cellular Homeostasis Is Altered in an RTT Mouse Model and Can Be Partially Reestablished by Exposure to a Neuronal Plasticity-Dependent Paradigm
It has been demonstrated that reactivation of Mecp2 neuronal expression can rescue the RTT phenotype in mice [51], supporting the idea that Mecp2 deficiency in neurons is sufficient to cause a RTT phenotype. However, in vitro evidence indicates that Mecp2 deficiency in glia may also contribute to brain dysfunction. Mecp2-deficient microglia cause dendritic and synaptic damage mediated by elevated glutamate (Glu) release [52]. Mecp2 is involved in Glu clearance through the regulation of Glu transporters (GLAST/EAAT1 and GLT-1/EAAT2), and Glutamine Synthetase in astrocytes; Glu clearance and production are abnormal in Mecp2-deficient astrocytes, probably contributing to the pathological process of RTT [53]. As mRNA expression of GLAST and GLT-1 glutamate transporters is decreased in Mecp2-null astrocytes [53], here we evaluated whether EE exposure can reestablish GLAST and GLT-1 mRNA expression in Mecp2-/y mice. The results show that EE exposure failed to restore mRNA levels of GLAST (Figure 4A) and GLT-1 (Figure 4B) to WT levels. When we evaluated protein levels of GLAST and GLT-1 in WT and Mecp2-/y mice exposed to SC (Figure 4C-D) and Mecp2-/y mice exposed to SC or EE (Figure 4E-F), we found that contrary to what was expected, GLAST and GLT-1 protein levels are increased in Mecp2-/y mice exposed to SC (Figure 4C-D). Then, we evaluated GLAST and GLT-1 protein levels in Mecp2-/y mice exposed to SC or EE and found that EE exposure decreased GLAST and GLT-1 protein levels in Mecp2-/y mice (Figure 4E-F). These results indicate that in Mecp2-/y mice, exposure to EE decreases the elevated levels of GLAST and GLT1 by a mechanism not associated with the control transcription of those coding genes.
The proper control of the motor cortex involves glutamatergic neurotransmission. Alternative splicing in the extracellular ligand binding domain of the AMPA receptors (AMPAR) generates two variants, i.e., flip and flop. The flop variant of the Gria2 gene, coding for GluA2 subunit of AMPA receptor, promotes the channel to close more rapidly, thus desensitizing the channel at a faster rate than the flip sequence [54]. It has been previously reported that the loss of Mecp2 affects flip/flop splicing of AMPAR genes, leading to a significant splicing shift in the flip/flop exon toward the flop inclusion [55]. We evaluated the flip/flop ratio of Gria1, Gria2, and Gria3 genes that encode GluA1, GluA2, and GluA3 AMPAR subunits, respectively. As expected, we found a significant decrease in Gria1 and Gria2 flip/flop ratio in Mecp2-/y mice exposed to SC, which was counteracted by exposure to EE (Figure 4G).
To get insight into molecular mechanisms that could be altered in the forebrain of Mecp2-/y mice, a single-replicate microarray analysis of forebrain samples from Mecp2-/y mice versus WT mice was performed to seek potential candidate genes whose expression could be re-established by EE exposure. The complete list of differentially expressed genes is shown in additional file 1. We performed an immune-related gene ontology (GO) enrichment analysis for all differentially expressed genes in the Mecp2-/y mice when compared with WT mice. We found the following enriched terms mainly upregulated in the Mecp2-/y mouse: “Toll-like receptor 4 signaling pathway” (GO:0034142), “Response to type I interferon” (GO:0034340), and “Cellular response to type I interferon” (GO:0071357), with Irak1 belonging to these inflammatory GO terms (Figure S1). As previously reported, we found that Irak1 mRNA levels were higher in the forebrain of Mecp2-/y mice. Increased aberrant expression of Irak1 has been shown to cause increased NF-kB activity in Mecp2-/y mice [56], and given its role in inflammation, we further evaluated Irak1 mRNA expression in WT mice exposed to SC using RT-qPCR, and Mecp2-/y mice both exposed to SC and EE (Figure 4H). As expected, Mecp2-/y mice exposed to SC showed increased Irak1 expression. However, Irak1 expression in Mecp2-/y mice exposed to EE did not differ from WT mice exposed to SC. These results show that the exposure to EE attenuates the molecular phenotype of Mecp2-/y mice associated with neurotransmission in the forebrain such as Glu levels, Gria splicing, and inflammation process, thus might be underlying the attenuated progression of the motor phenotype.
3.5. The Exposure to a Neuronal Plasticity-Dependent Paradigm Ameliorates Altered Distribution of Neuromuscular Synapses in an RTT Mouse Model
Respiratory impairment, including hyperventilation, periods of breath-holding, forced breathing, and apneas is one of the leading causes of premature death in RTT patients [6,8]. An attractive possibility to explain the increased lifespan and enhanced motor capacity observed in response to EE is improved neuromuscular communication leading to enhanced skeletal muscle activity. Thus, we next evaluated the distribution of neuromuscular synapses. To this aim, we used the diaphragm, a key respiratory muscle that contracts in response to phrenic nerve activity to allow breathing.
In SC, the diaphragm of Mecp2-/y mice exhibited wider endplate distribution compared to WT mice (Figure 5A and C), a phenotype associated to defects on neuromuscular junction assembly [57]. Remarkably, while EE exposure did not affect neuromuscular junction distribution in the diaphragm of WT mice, Mecp2-/y mice exposed to EE exhibited a rescued phenotype upon EE exposure (Figure 5B and C). Indeed, quantification shows that the relative abundance of wider endplates (>800 mm width) was significantly reduced after EE exposure in Mecp2-/y mice to reach values comparable to WT mice (Figure 5D). These findings demonstrate that a neuronal plasticity-dependent paradigm can induce changes in the distribution of the peripheral neuromuscular synapse. These changes likely contribute to the improved neurological and motor phenotypes and the increased lifespan observed in RTT mice in response to EE.
3.6. The Treatment with Young Plasma Attenuates the RTT-Like Phenotype in Mice
Our results showed that exposure to a neuronal plasticity-dependent paradigm induces neurological and motor improvement in an RTT mouse model associated with central and peripheral structural changes in neurons and at the neuromuscular junction. One of the components of our experimental paradigm is motor activity, which is reduced in Mecp2-/y mice. Besides, it has been demonstrated that physical activity increases the plasma level of neurotrophic factors [58] and that treatment with plasma from young to aged mice increases neuronal plasticity [45]. Thus, we hypothesize that the increased physical activity induced by EE in Mecp2-/y mice could modify plasma components with a neurological effect. Therefore, treatment with young plasma from WT mice with a similar physical activity level to Mecp2-/y mice could, at least in part, emulate the beneficial effect of exposure to an EE.
To test whether treatment with young plasma may decrease the neurological progression and behavioral phenotype exhibited by an RTT mouse model, the IP plasma injection started at 4 weeks of age, when mice exhibit a stronger phenotype compared to that at 3 weeks of age. As with EE, IP plasma injection increases Mecp2-/y mice lifespan (Figure 6A) and attenuates the motor and neurological RTT-like symptoms progression as weekly evaluated in the ledge test (Figure 6B) and hindlimb clasping (Figure 6C). To determine whether IP plasma injection recapitulates the elevated plus maze phenotype, we compared the place preference among WT, Mecp2-/y control mice, and those treated with IP plasma transference. As described above, Mecp2-/y mice stay longer in the open arm of the maze in comparison to their WT littermate, a phenotype that was not prevented by the IP young mice plasma transfer treatment (Figure 6D). However, as well as for the exposure to EE, the motor coordination phenotype exhibited by Mecp2-/y mice was prevented by IP young mice plasma injection. Similar to that described above, Mecp2-/y mice showed an increased time of the first arrival in the dowel test an increased number of falls. The IP young mice plasma injection prevents both phenotypes, reducing the time for the first arrival and the number of falls (Figure 6E and F). These results showed that the treatment with plasma from young WT mice, who have regular locomotor activity, prevents the motor phenotype of Mecp2-/y mice.
3.7. The Treatment with Young Plasma Decreases the Synaptic Deficit Exhibited by an RTT Mouse Model
To determine whether the IP injection of young mice plasma impacted the brain and neuronal cytoarchitecture, we performed Golgi staining in the brain from Mecp2-/y control mice and IP plasma-treated and were compared to their WT littermates (Figure 7A). A systemic therapeutical intervention such as the IP young mice plasma injection impacts the brain and neuronal cytoarchitecture. We observed that the diminished brain architecture observed in Mecp2-/y mice in the control group was prevented by the IP plasma injection, almost reestablishing the thickness of the corpus callosum (Figure 7B) and motor cortex (Figure 7C) like that observed in the WT littermate. Besides, as described above, the motor cortex of Mecp2-/y mice showed signs of moderate astrogliosis, which was attenuated by IP plasma injection (Figure 7D). Moreover, the neuronal cytoarchitecture of pyramidal neurons of Mecp2-/y mice was reestablished by IP plasma injections (Figure 7E), as evaluated by dendritic length (Figure 7F) and dendritic arborization complexity (Figure 7G). All these results show that the evaluated neurological parameters reduced in Mecp2-/y mice are prevented by the treatment of Mecp2-/y mice with IP young plasma injections.
4. Discussion
Environmental enrichment (EE) has been widely used to ameliorate the impaired neuronal functions in different rodent models of brain disorders [36,37,59], being a potential alternative or complement to the pharmacological treatment of neurodevelopmental disorders. In this work, we show how EE and treatment with young plasma decrease the progression of the Rett Syndrome phenotype in a mouse model of the disease. First, we evaluate the contribution of EE in the phenotype exhibited by Mecp2-/y mice. Body weight and lifespan were ameliorated by EE exposure since weaning. These results show that exposure to EE might be impacting the energy balance of mice lacking the expression of Mecp2. The role of Mecp2 as a master regulator of body weight has been explored by our and other’s labs [47,50,60,61] and its expression in Mecp2-/y mice reestablishes the increased body weight balance and increases lifespan [42]. However, the above effect of EE is independent of the expression of Mecp2, indicating that exposure to EE could be activating mechanisms downstream of Mecp2 expression that could partly compensate for its effect.
Further evaluation of RTT phenotype showed that parameters like corporal tremor, piloerection, and hind-limb discoordination that were increased in Mecp2-/y mice, were prevented by early EE exposure and the overall phenotypic evaluation of Mecp2-/y mice, measured as total score, showed that permanent exposure to EE since weaning attenuates most of the RTT phenotype. However, EE exposure was ineffective in preventing the increased clasping exhibited by Mecp2-/y mice, indicating that not all neurological functions are targeted by the EE downstream mechanism. On the other side, behavioral phenotypes like hypoactivity and abnormalities in locomotion, stereotypies, and anxiety reminiscent of the clinical condition have been reported in Mecp2 mouse mutants [17,50,62,63]. Here, we performed a battery of behavioral tests to determine whether EE exposure improves the clinical-like manifestation already reported. We and others have previously reported that Mecp2-/y mice exhibited behaviors that correlate with reduced anxiety in the elevated plus maze assay [42,64]. As expected, Mecp2-/y mice exposed to SC spend more time in the open arms, with no preference for the closed arms as WT mice, and EE exposure reestablished the preference for closed arms in Mecp2-/y mice. It is still known whether this effect of EE is related to a re-establishing of anxiety-associated behavior as a consequence of improving spatial perception. When we evaluated locomotion by measuring the distance traveled in an open field test, Mecp2-/y mice exposed to EE showed improved locomotion activity compared to Mecp2-/y mice exposed to SC. Results pointing in the same direction were found when motor control strength and coordination were evaluated, finding that EE exposure prevented the phenotype exhibited by Mecp2-/y mice. Mecp2-/y mice exposed to EE had fewer failures when hanging from the wire, fewer falls in the elevated dowel test, and improved latency time and the time of the first arrival compared to Mecp2-/y mice exposed to SC. Moreover, the number of arrivals was reestablished, exhibiting a performance comparable to that of WT mice. Motor coordination and learning were assessed on the rotating cylinder of a rotarod. Early EE exposure improved poor motor coordination in Mecp2-/y mice but failed to ameliorate motor learning. All these results showed that early exposure to EE positively impacts counteracting abnormalities in muscle strength, locomotor activity, and motor coordination. However, these results also show that the expression of Mecp2 is required for motor memory formation, as has been demonstrated for spatial learning in this mouse model [49].
To investigate further how EE improves locomotor activity and motor function in Mecp2-/y mice, we evaluated the phenotype of the motor cortex to determine whether EE exposure might modulate neuromotor progression by changes in neuronal cytoarchitecture as has been demonstrated in other rodent models [59]. We evaluated the M1-M2 motor cortex cytoarchitecture at 3 and 7 weeks of age, in WT and Mecp2-/y mice exposed to either housing SC or EE. At 3 weeks of age, the RTT phenotype is just starting to appear, whereas at 7 weeks of age, Mecp2-/y mice showed severe neurological RTT-like symptoms [15]. Also, it was the time at which we observed motor performance improvement in response to EE exposure. As expected, the motor cortex of 3 weeks of age Mecp2-/y mice is not far different from WT. However, at 7 weeks of age, the motor cortex of Mecp2-/y mice is disorganized, with reduced neuronal complexity and signs of moderate astrogliosis. After 4 weeks of EE exposure, the motor cortex of Mecp2-/y mice was better organized, had improved neuronal complexity, and had no signs of astrogliosis, seeming similar to WT mice. A more detailed evaluation of pyramidal neurons of cortical layer V was performed, where we measured the dendritic length and the number of dendrites in pyramidal neurons. Mecp2-/y mice housed in SC have a reduction in the dendritic length and the number of dendrites compared to WT mice. However, these differences were not observed in the pyramidal neurons of Mecp2-/y exposed to EE. Additionally, second-order dendrites of pyramidal neurons from the motor cortex of Mecp2-/y mice have less spine density compared to WT, but when exposed to EE, the spine density in Mecp2-/y mice was both higher than Mecp2-/y mice in SC and similar to WT. Thus, these results show that early exposure to an EE that increased neuronal plasticity and has a positive impact on preventing the damage in motor cortex cytoarchitecture associated with RTT neurological progression. To get insight into the mechanism by which EE exposure could be avoiding the deterioration of the motor cortex, we next evaluate molecular parameters associated with neurotransmission.
Rett phenotype has been initially associated with a neuronal Mecp2 loss of function [16]. There is growing evidence that Mecp2 deficiency in glia contributes to brain dysfunction and, therefore, to RTT progression [52,53]. Glutamate clearance and production, processes controlled by Glu transporters and Glutamine Synthetase, are abnormal in Mecp2-deficient astrocytes in vitro, probably contributing to the pathological process of RTT [53], and mRNA expression of GLAST and GLT-1 glutamate transporters is decreased in Mecp2-null astrocytes [53]. Thus, here we evaluated whether the exposure to EE restored mRNA levels of GLAST and GLT-1 to WT levels in Mecp2-/y mice, and surprisingly, we found no effect of EE exposure on GLAST nor GLT1 mRNA expression. Strikingly, we found an increase in GLAST and GLT-1 protein levels in Mecp2-/y mice exposed to SC compared to WT mice, and EE exposure decreased GLAST and GLT-1 protein levels in this mouse model of RTT. Some studies have demonstrated increased Glu levels in the cerebrospinal fluid (CSF) of RTT patients [65,66]. Moreover, Mecp2-deficient microglia release a high level of Glu [52], involving Mecp2 in modulating Glu metabolism. With these antecedents, we did not expect to find increased levels of GLAST and GLT-1 transporters in Mecp2-/y mice since this could increase the uptake of Glu, reducing its levels. However, we do not know the localization of these transporters; whether they are in the cell surface is unknown; thus, further experiments are required to answer this and other questions regarding Glu metabolism in Mecp2-/y mice exposed to EE. Another parameter related to synaptic activity is glutamate receptors. AMPA receptors (AMPAR) subunits are alternatively spliced, generating the “flip” and “flop” variants, which have different kinetic properties [54]. Interestingly, the loss of Mecp2 affects flip/flop splicing of AMPAR genes, leading to a significant splicing shift to the flop inclusion, leading to a faster decay of AMPAR-gated current, and altered synaptic transmission [55], which could impact NMDA receptor activity and compromise synaptic transmission. As expected, we found a significant decrease in Gria1 and Gria2 flip/flop ratio in Mecp2-/y mice exposed to SC, and according to the behavioral results described above, EE exposure reestablished the flip/flop ratio to WT levels. These results together indicate that the effect of EE exposure attenuating the RTT phenotype and the RTT-like behavior might be mediated by improving neuronal function through reestablishing glutamatergic homeostasis and neurotransmission. However, further analysis must be performed to evaluate electrical synaptic properties.
One of the mechanisms underlying defective neurotransmission is neuroinflammation [67,68]. Irak1 mRNA expression is upregulated in Mecp2-/y mice, leading to an increased NF-kB signaling [56], as expected, Mecp2-/y mice exposed to SC showed an increase in Irak1 expression, and EE exposure decreased Irak1 mRNA expression to similar levels as in WT mice. NF-kB signaling is becoming increasingly recognized as a regulator of the growth and morphology of neural processes in the developing and mature nervous system [69]. Therefore, altered NF-kB signaling could play a significant role in RTT progression [56]. Moreover, there is evidence showing that Irak1 gene is duplicated in patients with the MECP2 duplication syndrome [70] and that drugs targeting Irak1 in vitro rescue the inflammatory phenotype associated with RTT [71]. Moreover, the Golgi staining analysis of Mecp2-/y mice exposed to EE is coherent with the expected phenotype associated with a decreased neuroinflammation. Thus, although the effect of EE on Irak1 protein levels was not evaluated, our results get some light on the potential use of EE-based strategies to attenuate Irak1 expression as a novel non-pharmacological therapeutic strategy to modulate NF-kB signaling to reduce neuroinflammation in RTT patients.
Respiratory impairment is one of the leading premature causes of death in RTT patients [6,8]. In RTT mouse models, irregular breathing and hard respiration have been reported [15,16]. The phrenic-diaphragm neuromuscular junction in Mecp2-/y mice had an increased endplate compared with their WT littermates, and we found that this increase was prevented by exposure to EE in Mecp2-/y mice. Thus, exposure to a neuronal plasticity-dependent paradigm induces changes in the neuromuscular junction that could be underlying the improved neurological, motor phenotype, and lifespan observed in response to this experimental paradigm. This improved neuromuscular junction induced by EE exposure could be extensive to skeletal muscles, improving motor function and the locomotor activity associated with EE effect on Mecp2-/y mice; therefore, increasing the release of neurotrophic myokines that could be involved in attenuating the progression of RTT phenotype in this mouse model. As an approach to test this hypothesis, we evaluated the effect of the treatment with plasma from young mice with regular motor activity on the progression of RTT phenotype.
It has been previously demonstrated that plasma treatment from young to aged mice increases neuronal plasticity [45], probably by delivering increased levels of neurotrophic factors like BDNF [58], which circulating levels are decreased in both RTT patients and Mecp2-/y mice. Therefore, young plasma treatment from WT mice with regular physical activity could replicate some beneficial effects observed in mice exposed to EE. After 2,5 weeks of young plasma IP injections, Mecp2-/y mice had attenuated motor and neurological RTT-like symptoms progression, evaluated by ledge test and hindlimb clasping, and over time, increased lifespan. However, IP young plasma injections did not prevent open arms preference in elevated plus maze observed in Mecp2-/y mice. Still, it did improve poor motor coordination in Mecp2-/y mice, as the time of first arrival and the number of falls from the elevated dowel test were similar to WT mice. Young plasma injections almost reestablished the thickness of the corpus callosum and motor cortex of Mecp2-/y mice to that in WT mice, preventing the diminished brain architecture observed in Mecp2-/y mice in SC. The moderate astrogliosis presented in the motor cortex of Mecp2-/y mice was attenuated by IP plasma injection, and the neuronal cytoarchitecture of pyramidal neurons evaluated by dendritic length and dendritic arborization complexity was re-established by the plasma treatment. Hence, our results not only show the potential therapeutic effect of plasma treatment to attenuate RTT phenotype, but also show that peripheral signals are conceivable to reactivate the dormant neurodevelopment in RTT.
Currently, there is no cure for RTT syndrome; several therapeutic avenues have been explored; the most recent is trofinetide, an FDA-approved drug; despite its therapeutic potential, adverse side effects have been reported, including diarrhea, seizures, and vomiting [72]. Thus, finding treatment alternatives with few to no adverse side effects like the ones presented here is significant in improving the quality of life and symptoms of RTT patients.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Differentially expressed genes between Mecp2-/y and wild type mice are enriched in genes belonging to Immune System Processes terms and pathways. The Differentially expressed genes list between Mecp2-/y and wild type mice (see Additional File 1) was used as input for ClueGO app in Cytoscape for Gene Ontology Enrichment Analysis using GO ImmuneSystemProcess-EBI-UniProt-GOA-ACAP-ARAP_25.05.2022_00h00 : 3113 as reference database. Default parameters were used to create the ClueGO network. The node size represented the significance (small node represented p > 0.1, medium node represented 0.05 > p > 0.0005, and big node represented p < 0.0005). The different colors on each node represented the specificity of the biological role of the genes. Genes associated with antigen processing and presentation of peptide antigen via MHC class Ib, positive regulation of pattern recognition receptor signaling pathway, toll-like receptor 4 signaling pathway, antigen processing and presentation of endogenous peptide antigen via MHC class Ib, antigen processing and presentation of endogenous peptide antigen, response to type I interferon, and cellular response to type I interferon.
Author Contributions
Conceptualization, CN and BK; Methodology, CN, RT, NLl, VV, CS, AV, EN-P, JS-C, JM, RK. SV, MP-B, PO-P, and NP.; software, DL, SE; validation, SE and BK; formal analysis, SE and JM; investigation, CN, SE, SB, JPH, BK.; resources, BK, RT and SE.; data curation, BK and SE; writing—original draft preparation, SE and BK.; writing—review and editing, BK, JPH and SE; funding acquisition, SE, RT, JPH, and BK. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by Fondecyt 1181574, 1230905 and Anillo ACT210039 (BK), Fondecyt 3180518 and 11230898 (RT), Fondecyt Postdoctorado 3210493 (SE), Fondecyt 1221213 (JPH).
Institutional Review Board Statement
“The animal study protocol was approved by the Institutional Ethics Committee of Centro de Estudios Científicos-CECs (IACUC XXX 2015) and (IACUC XXX 2016).” for studies involving animals.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rett, A. Uber Ein Eigenartiges Hirnatrophisches Syndrom Bei Hyperammonaemie in Kindesalter. Wien Med Wochenschr 1966, 116, 723–738.
- Laurvick, C.L.; de Klerk, N.; Bower, C.; Christodoulou, J.; Ravine, D.; Ellaway, C.; Williamson, S.; Leonard, H. Rett Syndrome in Australia: A Review of the Epidemiology. J. Pediatr 2006, 148, 347–352. [CrossRef]
- Amir, R.E.; Van Den Veyver, I.B.; Wan, M.; Tran, C.Q.; Francke, U.; Huda, &; Zoghbi, Y. Rett Syndrome Is Caused by Mutations in X-Linked MECP2, Encoding Methyl-CpG-Binding Protein 2. Nat Genet 1999, 23, 185–188. [CrossRef]
- Chahrour, M.; Zoghbi, H.Y. The Story of Rett Syndrome: From Clinic to Neurobiology. Neuron 2007, 56, 422–437. [CrossRef]
- Liyanage, V.R.B.; Rastegar, M. Rett Syndrome and MeCP2. Neuromolecular Med 2014, 16, 231–264. [CrossRef]
- Vashi, N.; Justice, M.J. Treating Rett Syndrome: From Mouse Models to Human Therapies. Mammalian Genome 2019, 30, 90–110. [CrossRef]
- Jellinger, K.A. Neuropathology of Movement Disorders. Neurosurg Clin N Am 1998, 9, 237–262.
- Tarquinio, D.C.; Hou, W.; Neul, J.L.; Kaufmann, W.E.; Glaze, D.G.; Motil, K.J.; Skinner, S.A.; Lee, H.S.; Percy, A.K. The Changing Face of Survival in Rett Syndrome and MECP2-Related Disorders. Pediatr Neurol 2015, 53, 402–411. [CrossRef]
- Amir, R.E.; Zoghbi, H.Y. Rett Syndrome: Methyl-CpG-Binding Protein 2 Mutations and Phenotype-Genotype Correlations. Am J Med Genet . 2000, 97, 147–152. [CrossRef]
- Wan, M.; Lee, S.S.J.; Zhang, X.; Houwink-Manville, I.; Song, H.R.; Amir, R.E.; Budden, S.; Naidu, S.B.; Pereira, J.L.P.; Lo, I.F.M.; et al. Rett Syndrome and beyond: Recurrent Spontaneous and Familial MECP2 Mutations at CpG Hotspots. Am J Hum Genet 1999, 65, 1520–1529. [CrossRef]
- Meehan, R.R.; Lewis, J.D.; McKay, S.; Kleiner, E.L.; Bird, A.P. Identification of a Mammalian Protein That Binds Specifically to DNA Containing Methylated CpGs. Cell 1989, 58, 499–507. [CrossRef]
- Chahrour, M.; Jung, S.Y.; Shaw, C.; Zhou, X.; Wong, S.T.C.; Qin, J.; Zoghbi, H.Y. MeCP2, a Key Contributor to Neurological Disease, Activates and Represses Transcription. Science (1979) 2008, 320, 1224–1229. [CrossRef]
- Shahbazian MD; Antalffy B; Armstrong DL; Zoghbi HY Insight into Rett Syndrome: MeCP2 Levels Display Tissue- and Cell-Specific Differences and Correlate with Neuronal Maturation. Hum Mol Genet 2002, 11, 115–124. [CrossRef]
- Bedogni F; Cobolli Gigli C; Pozzi D; Rossi RL; Scaramuzza L; Rossetti G; Pagani M; Kilstrup-Nielsen C; Matteoli M; Landsberger N Defects During Mecp2 Null Embryonic Cortex Development Precede the Onset of Overt Neurological Symptoms. Cereb Cortex 2016, 26, 2517–2529. [CrossRef]
- Guy J; Hendrich B; Holmes M; Martin JE; Bird A A Mouse Mecp2-Null Mutation Causes Neurological Symptoms That Mimic Rett Syndrome. Nat Genet. 2001, 27, 322–326. [CrossRef]
- Chen RZ; Akbarian S; Tudor M; Jaenisch R Deficiency of Methyl-CpG Binding Protein-2 in CNS Neurons Results in a Rett-like Phenotype in Mice. Nat. Genet 2001, 27, 327–331. [CrossRef]
- Pelka GJ; Watson CM; Radziewic T; Hayward M; Lahooti H; Christodoulou J; Tam PP Mecp2 Deficiency Is Associated with Learning and Cognitive Deficits and Altered Gene Activity in the Hippocampal Region of Mice. Brain 2006, 129, 887–898. [CrossRef]
- Kyle, S.M.; Vashi, N.; Justice, M.J. Rett Syndrome: A Neurological Disorder with Metabolic Components. Open Biol 2018, 8. [CrossRef]
- Lombardi, L.M.; Baker, S.A.; Zoghbi, H.Y. MECP2 Disorders: From the Clinic to Mice and Back. Journal of Clinical Investigation 2015, 125, 2914–2923. [CrossRef]
- Nguyen, M.V.C.; Du, F.; Felice, C.A.; Shan, X.; Nigam, A.; Mandel, G.; Robinson, J.K.; Ballas, N. MeCP2 Is Critical for Maintaining Mature Neuronal Networks and Global Brain Anatomy during Late Stages of Postnatal Brain Development and in the Mature Adult Brain. Journal of Neuroscience 2012, 32, 10021–10034. [CrossRef]
- Derecki, N.C.; Cronk, J.C.; Lu, Z.; Xu, E.; Abbott, S.B.G.; Guyenet, P.G.; Kipnis, J. Wild-Type Microglia Arrest Pathology in a Mouse Model of Rett Syndrome. Nature 2012, 484, 105–109. [CrossRef]
- Giacometti, E.; Luikenhuis, S.; Beard, C.; Jaenisch, R. Partial Rescue of MeCP2 Deficiency by Postnatal Activation of MeCP2. PNAS 2007, 104, 1931–1936. [CrossRef]
- Guy, J.; Gan, J.; Selfridge, J.; Cobb, S.; Bird, A. Reversal of Neurological Defects in a Mouse Model of Rett Syndrome. Science (1979) 2007, 315, 1143–1147. [CrossRef]
- Ballas, N.; Lioy, D.T.; Grunseich, C.; Mandel, G. Non-Cell Autonomous Influence of MeCP2-Deficient Glia on Neuronal Dendritic Morphology. Nat Neurosci 2009, 12, 311–317. [CrossRef]
- Xu, X.; Miller, E.C.; Pozzo-Miller, L. Dendritic Spine Dysgenesis in Rett Syndrome. Front Neuroanat 2014, 8, 1–8. [CrossRef]
- Belichenko PV; Oldfors A; Hagberg B; Dahlström A Rett Syndrome: 3-D Confocal Microscopy of Cortical Pyramidal Dendrites and Afferents; 1994; Vol. 5, pp. 1509–1513;.
- Armstrong DD The Neuropathology of Rett Syndrome--Overview 1994. Neuropediatrics 1995, 26, 100–104. [CrossRef]
- Chapleau, C.A.; Calfa, G.D.; Lane, M.C.; Albertson, A.J.; Larimore, J.L.; Kudo, S.; Armstrong, D.L.; Percy, A.K.; Pozzo-Miller, L. Dendritic Spine Pathologies in Hippocampal Pyramidal Neurons from Rett Syndrome Brain and after Expression of Rett-Associated MECP2 Mutations. Neurobiol Dis 2009, 35, 219–233. [CrossRef]
- Kim, K.Y.; Hysolli, E.; Park, I.H. Neuronal Maturation Defect in Induced Pluripotent Stem Cells from Patients with Rett Syndrome. Proc Natl Acad Sci U S A 2011, 108, 14169–14174. [CrossRef]
- Patriarchi, T.; Amabile, S.; Frullanti, E.; Landucci, E.; Lo Rizzo, C.; Ariani, F.; Costa, M.; Olimpico, F.; Hell, J.W.; Vaccarino, F.M.; et al. Imbalance of Excitatory/Inhibitory Synaptic Protein Expression in IPSC-Derived Neurons from FOXG1 Patients and in Foxg1 Mice. European Journal of Human Genetics 2016, 24, 871–880. [CrossRef]
- Belichenko NP; Belichenko PV; Mobley WC. Evidence for Both Neuronal Cell Autonomous and Nonautonomous Effects of Methyl-CpG-Binding Protein 2 in the Cerebral Cortex of Female Mice with Mecp2 Mutation. Neurobiol Dis 2009, 34, 71–77. [CrossRef]
- Fukuda T; Itoh M; Ichikawa T; Washiyama K; Goto Y Delayed Maturation of Neuronal Architecture and Synaptogenesis in Cerebral Cortex of Mecp2-Deficient Mice. J Neuropathol Exp Neurol 2005, 64, 537–544. [CrossRef]
- Stuss, D.; Boyd, J.; Levin, D.; Delaney, K. MeCP2 Mutation Results in Compartment-Specific Reductions in Dendritic Branching and Spine Density in Layer 5 Motor Cortical Neurons of YFP-H Mice. PLoS One 2012, 7. [CrossRef]
- Landi S; Putignano E; Boggio EM; Giustetto M; Pizzorusso T; Ratto GM The Short-Time Structural Plasticity of Dendritic Spines Is Altered in a Model of Rett Syndrome. Sci Rep 2011, 1. [CrossRef]
- Blackman, M.P.; Djukic, B.; Nelson, S.B.; Turrigiano, G.G. A Critical and Cell-Autonomous Role for MeCP2 in Synaptic Scaling Up. Journal of Neuroscience 2012, 32, 13529–13536. [CrossRef]
- Nithianantharajah J; Hannan AJ Enriched Environments, Experience-Dependent Plasticity and Disorders of the Nervous System. Nat Rev Neurosci 2006, 7, 697–709. [CrossRef]
- Mohd Sahini SN; Mohd Nor Hazalin NA; Srikumar BN; Jayasingh Chellammal HS; Surindar Singh GK Environmental Enrichment Improves Cognitive Function, Learning, Memory and Anxiety-Related Behaviours in Rodent Models of Dementia: Implications for Future Study. Neurobiol Learn Mem 2024, 208. [CrossRef]
- Kondo, M.; Gray, L.; Pelka, G.; Christodoulou, J.; Tam, P.; Hannan, A. Environmental Enrichment Ameliorates a Motor Coordination Deficit in a Mouse Model of Rett Syndrome--Mecp2 Gene Dosage Effects and BDNF Expression. Eur J Neurosci 2008, 27, 3342–3350. [CrossRef]
- Nag N; Moriuchi JM; Peitzman CG; Ward BC; Kolodny NH; Berger-Sweeney JE Environmental Enrichment Alters Locomotor Behaviour and Ventricular Volume in Mecp2 1lox Mice. Behav Brain Res 2009, 3, 44–48. [CrossRef]
- Kerr, B.; Silva, P.; Walz, K.; Young, J. Unconventional Transcriptional Response to Environmental Enrichment in a Mouse Model of Rett Syndrome. PLoS One 2010, 5. [CrossRef]
- Lonetti, G.; Angelucci, A.; Morando, L.; Boggio, E.; Giustetto, M.; Pizzorusso, T. Early Environmental Enrichment Moderates the Behavioral and Synaptic Phenotype of MeCP2 Null Mice. Biol Psychiatry 2010, 67, 657–665. [CrossRef]
- Kerr, B.; Soto C, J.; Saez, M.; Abrams, A.; Walz, K.; Young, J.I. Transgenic Complementation of MeCP2 Deficiency: Phenotypic Rescue of Mecp2-Null Mice by Isoform-Specific Transgenes. European Journal of Human Genetics 2012, 20, 69–76. [CrossRef]
- Gelfo, F.; Cutuli, D.; Foti, F.; Laricchiuta, D.; De-Bartolo, P.; Caltagirone, C.; Petrosini, L.; Angelucci, F. Enriched Environment Improves Motor Function and Increases Neurotrophins in Hemicerebellar Lesioned Rats. Neurorehabil Neural Repair 2011, 25, 243–252. [CrossRef]
- Turner, C.A.; Lewis, M.H. Environmental Enrichment: Effects on Stereotyped Behavior and Neurotrophin Levels. Physiol Behav 2003, 80, 259–266. [CrossRef]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young Blood Reverses Age-Related Impairments in Cognitive Function and Synaptic Plasticity in Mice. Nat Med 2014, 20, 659–663. [CrossRef]
- Torres, R.F.; Hidalgo, C.; Kerr, B. Mecp2 Mediates Experience-Dependent Transcriptional Upregulation of Ryanodine Receptor Type-3. Front Mol Neurosci 2017, 10. [CrossRef]
- Torres-Andrade, R.; Moldenhauer, R.; Gutierrez-Bertín, N.; Soto-Covasich, J.; Mancilla-Medina, C.; Ehrenfeld, C.; Kerr, B. The Increase in Body Weight Induced by Lack of Methyl CpG Binding Protein-2 Is Associated with Altered Leptin Signalling in the Hypothalamus. Exp Physiol 2014, 99, 1229–1240. [CrossRef]
- Schmittgen TD; Livak KJ Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat Protoc. 2008, 3, 1101–1108. [CrossRef]
- Torres, R.F.; Hidalgo, C.; Kerr, B. Mecp2 Mediates Experience-Dependent Transcriptional Upregulation of Ryanodine Receptor Type-3. Front Mol Neurosci 2017, 10. [CrossRef]
- Kerr, B.; Alvarez-saavedra, M.; Sáez, M.A.; Saona, A.; Young, J.I. Defective Body-Weight Regulation, Motor Control and Abnormal Social Interactions in Mecp2 Hypomorphic Mice. Hum Mol Genet 2008, 17, 1707–1717. [CrossRef]
- Calfa, G.; Percy, A.K.; Pozzo-Miller, L. Experimental Models of Rett Syndrome Based on Mecp2 Dysfunction. Exp Biol Med 2011, 236, 3–19. [CrossRef]
- Maezawa, I.; Jin, L.W. Rett Syndrome Microglia Damage Dendrites and Synapses by the Elevated Release of Glutamate. Journal of Neuroscience 2010, 30, 5346–5356. [CrossRef]
- Okabe, Y.; Takahashi, T.; Mitsumasu, C.; Kosai, K. ichiro; Tanaka, E.; Matsuishi, T. Alterations of Gene Expression and Glutamate Clearance in Astrocytes Derived from an Mecp2-Null Mouse Model of Rett Syndrome. PLoS One 2012, 7. [CrossRef]
- Pei W; Huang Z; Wang C; Han Y; Park JS; Niu L Flip and Flop: A Molecular Determinant for AMPA Receptor Channel Opening. Biochemistry 2009, 48, 3767–3777. [CrossRef]
- Li, R.; Dong, Q.; Yuan, X.; Zeng, X.; Gao, Y.; Chiao, C.; Li, H.; Zhao, X.; Keles, S.; Wang, Z.; et al. Misregulation of Alternative Splicing in a Mouse Model of Rett Syndrome. PLoS Genet 2016, 12. [CrossRef]
- Kishi, N.; MacDonald, J.L.; Ye, J.; Molyneaux, B.J.; Azim, E.; Macklis, J.D. Reduction of Aberrant NF-ΚB Signalling Ameliorates Rett Syndrome Phenotypes in Mecp2-Null Mice. Nat Commun 2016, 7. [CrossRef]
- Henriquez, J.P.; Webb, A.; Bence, M.; Bildsoe, H.; Sahores, M.; Hughes, S.M.; Salinas, P.C. Wnt Signaling Promotes AChR Aggregation at the Neuromuscular Synapse in Collaboration with Agrin; 2008;
- Phillips, C. Brain-Derived Neurotrophic Factor, Depression, and Physical Activity: Making the Neuroplastic Connection. Neural Plast 2017, 2017. [CrossRef]
- Hannan, A.J. Review: Environmental Enrichment and Brain Repair: Harnessing the Therapeutic Effects of Cognitive Stimulation and Physical Activity to Enhance Experience-Dependent Plasticity. Neuropathol Appl Neurobiol 2014, 40, 13–25. [CrossRef]
- Nuria Llontop, A.; Mancilla, C.; Ojeda-Provoste, P.; Torres, A.K.; Godoy, A.; Tapia-Rojas, C.; Kerr, B.; Tapia-Rojas Centro científico, C. The Methyl-CpG-Binding Protein 2 (Mecp2) Regulates the Hypothalamic Mitochondrial Function and White Adipose Tissue Lipid Metabolism;
- Wang, X.; Lacza, Z.; Sun, Y.E.; Han, W. Leptin Resistance and Obesity in Mice with Deletion of Methyl-CpG-Binding Protein 2 (MeCP2) in Hypothalamic pro-Opiomelanocortin (POMC) Neurons. Diabetologia 2014, 57, 236–245. [CrossRef]
- Mcgill, B.E.; Bundle, S.F.; Yaylaoglu, M.B.; Carson, J.P.; Thaller, C.; Zoghbi, H.Y. Enhanced Anxiety and Stress-Induced Corticosterone Release Are Associated with Increased Crh Expression in a Mouse Model of Rett Syndrome. 2006. [CrossRef]
- Stearns NA; Schaevitz LR; Bowling H; Nag N; Berger UV; Berger-Sweeney J Behavioral and Anatomical Abnormalities in Mecp2 Mutant Mice: A Model for Rett Syndrome. Neuroscience 2007, 146, 907–921. [CrossRef]
- Yasui, D.H.; Gonzales, M.L.; Aflatooni, J.O.; Crary, F.K.; Hu, D.J.; Gavino, B.J.; Golub, M.S.; Vincent, J.B.; Schanen, N.C.; Olson, C.O.; et al. Mice with an Isoform-Ablating Mecp2exon 1 Mutation Recapitulate the Neurologic Deficits of Rett Syndrome. Hum Mol Genet 2014, 23, 2447–2458. [CrossRef]
- Hamberger A; Gillberg C; Palm A; Hagberg B Elevated CSF Glutamate in Rett Syndrome. Neuropediatrics 1992, 23, 212–213. [CrossRef]
- Lappalainen R; Riikonen RS High Levels of Cerebrospinal Fluid Glutamate in Rett Syndrome. Pediatr Neurol 1996, 15, 213–216. [CrossRef]
- Mandolesi, G.; Gentile, A.; Musella, A.; Fresegna, D.; De Vito, F.; Bullitta, S.; Sepman, H.; Marfia, G.A.; Centonze, D. Synaptopathy Connects Inflammation and Neurodegeneration in Multiple Sclerosis. Nat Rev Neurol 2015, 11, 711–724. [CrossRef]
- Qi, C.; Guo, B.; Ren, K.; Yao, H.; Wang, M.; Sun, T.; Cai, G.; Liu, H.; Li, R.; Luo, C.; et al. Chronic Inflammatory Pain Decreases the Glutamate Vesicles in Presynaptic Terminals of the Nucleus Accumbens. Mol Pain 2018, 14, 1744806918781259. [CrossRef]
- Gutierrez, H.; Davies, A.M. Regulation of Neural Process Growth, Elaboration and Structural Plasticity by NF-ΚB. Trends Neurosci 2011, 34, 316–325.
- Peters, S.U.; Fu, C.; Suter, B.; Marsh, E.; Benke, T.A.; Skinner, S.A.; Lieberman, D.N.; Standridge, S.; Jones, M.; Beisang, A.; et al. Characterizing the Phenotypic Effect of Xq28 Duplication Size in MECP2 Duplication Syndrome. Clin Genet 2019, 95, 575–581. [CrossRef]
- Unterman, I.; Bloch, I.; Cazacu, S.; Kazimirsky, G.; Ben-Zeev, B.; Berman, B.P.; Brodie, C.; Tabach, Y. Expanding the MECP2 Network Using Comparative Genomics Reveals Potential Therapeutic Targets for Rett Syndrome. Elife 2021, 10, e67085. [CrossRef]
- Neul, J.L.; Percy, A.K.; Benke, T.A.; Berry-Kravis, E.M.; Glaze, D.G.; Marsh, E.D.; Lin, T.; Stankovic, S.; Bishop, K.M.; Youakim, J.M. Trofinetide for the Treatment of Rett Syndrome: A Randomized Phase 3 Study. Nat Med 2023, 29, 1468–1475. [CrossRef]
Figure 1.
Permanent exposure to an enriched environment (EE) since weaning attenuates the RTT-like phenotype in mice. (A) Body weight recordings of WT mice exposed to an enriched environment (EE) or standard cages (SC), and Mecp2-/y mice exposed to SC or EE, Mecp2-/y mice exposed to EE showed a decrease in body weight in comparison with Mecp2-/y mice housed in control conditions. (B) Lifespan of Mecp2-/y mice exposed to SC or EE. The decreased lifespan of Mecp2-/y mice is extended by exposure to EE. (C-G) Overall phenotypic evaluation of Mecp2-/y mice, (C) the increased level of clasping showed by Mecp2-/y mice was not prevented by exposing mice to EE. The increased levels of corporal tremor (D), piloerection (E), and hind-limb discoordination (F) showed by Mecp2-/y mice were prevented by exposure to EE. (G) General state evaluation of mice by a total score, which was higher in Mecp2-/y mice in SC compared to WT and diminished by exposure to EE in Mecp2-/y mice. These analyses were completed by (B) Simple survival analysis Mantel-Cox test followed by Wilcoxon test and (A, C-G) two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p <0.05, ✱✱p, ++p or ##p <0.01 and ✱✱✱✱p, ++++p or ####p<0.0001; ns, non-statistical differences.
Figure 1.
Permanent exposure to an enriched environment (EE) since weaning attenuates the RTT-like phenotype in mice. (A) Body weight recordings of WT mice exposed to an enriched environment (EE) or standard cages (SC), and Mecp2-/y mice exposed to SC or EE, Mecp2-/y mice exposed to EE showed a decrease in body weight in comparison with Mecp2-/y mice housed in control conditions. (B) Lifespan of Mecp2-/y mice exposed to SC or EE. The decreased lifespan of Mecp2-/y mice is extended by exposure to EE. (C-G) Overall phenotypic evaluation of Mecp2-/y mice, (C) the increased level of clasping showed by Mecp2-/y mice was not prevented by exposing mice to EE. The increased levels of corporal tremor (D), piloerection (E), and hind-limb discoordination (F) showed by Mecp2-/y mice were prevented by exposure to EE. (G) General state evaluation of mice by a total score, which was higher in Mecp2-/y mice in SC compared to WT and diminished by exposure to EE in Mecp2-/y mice. These analyses were completed by (B) Simple survival analysis Mantel-Cox test followed by Wilcoxon test and (A, C-G) two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p <0.05, ✱✱p, ++p or ##p <0.01 and ✱✱✱✱p, ++++p or ####p<0.0001; ns, non-statistical differences.
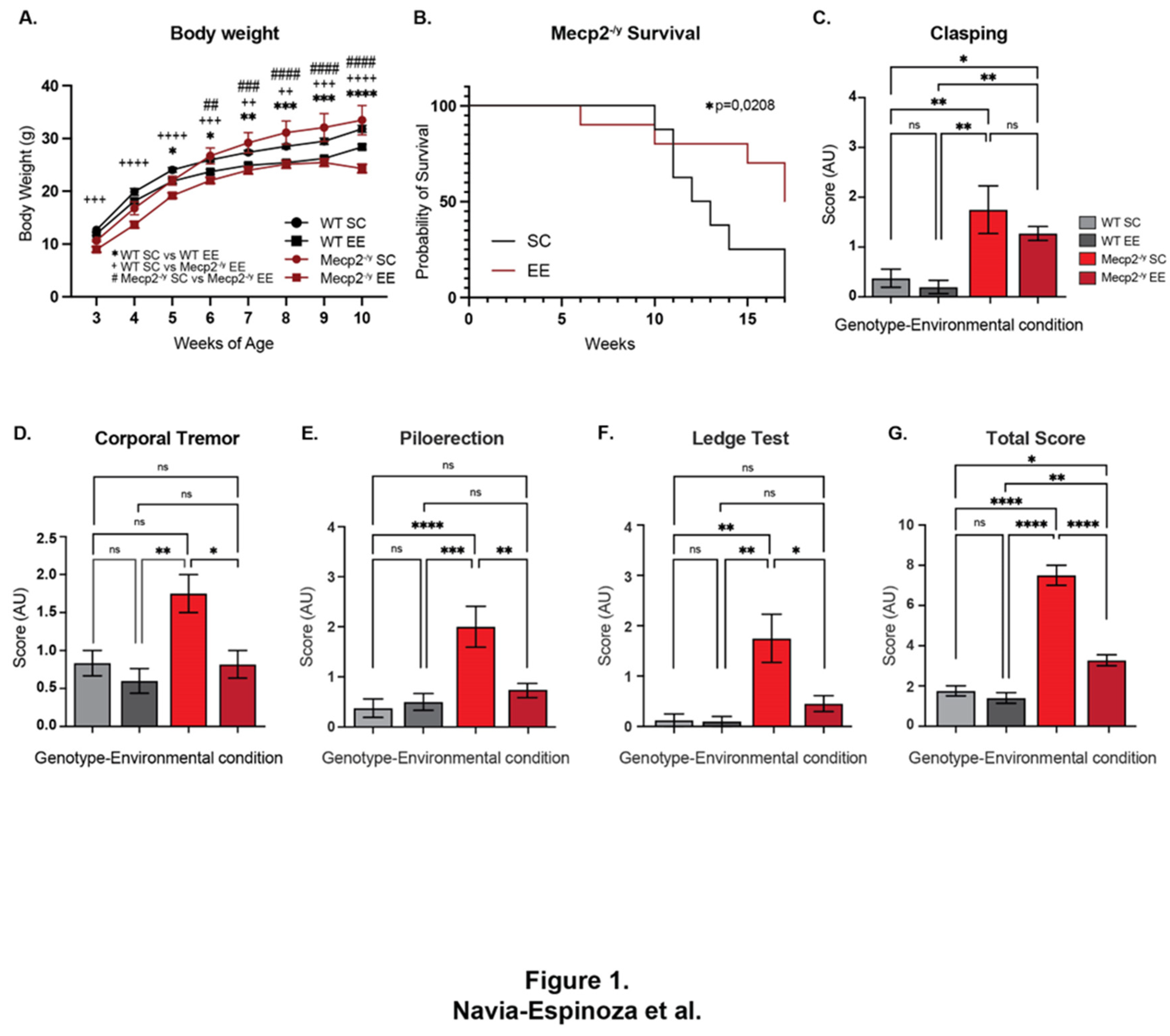
Figure 2.
Exposure to a neuronal plasticity-dependent paradigm to increase neuronal plasticity attenuates the behavioral phenotype and motor deficits in an RTT mouse model. (A) Elevated plus maze to evaluate anxiety-like behavior, the exposure to EE reestablished the preference for closed arms in Mecp2-/y mice. (B) Traveled distance in open field test, as a locomotion parameter, Mecp2-/y mice exposed to EE showed improved locomotion activity compared to Mecp2-/y mice exposed to SC. (C) Wire-hanging test to evaluate motor function, Mecp2-/y mice exposed to EE showed less failure in hanging from the wire in comparison with Mecp2-/y mice in SC. (D-G) Elevated dowel test to evaluate motor function, coordination and control, (D) Mecp2-/y mice in EE showed a similar number of falls as WT mice and fewer falls than Mecp2-/y mice in SC; EE exposure improved latency time (E) and the time of the first arrival (F) in Mecp2-/y mice, since they were similar to those exhibited by WT mice. (G) The number of arrivals of Mecp2-/y mice in SC were less than in WT mice, and this phenotype was completely prevented by the exposure to EE. (h) Motor coordination and learning was evaluated on the rotating cylinder of a rotarod, the poor motor coordination in Mecp2-/y mice were prevented by the exposure to EE, however motor learning was not recovered. These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p<0.05, ✱✱p <0.01, ✱✱✱p <0.001 and ✱✱✱✱p <0.0001.
Figure 2.
Exposure to a neuronal plasticity-dependent paradigm to increase neuronal plasticity attenuates the behavioral phenotype and motor deficits in an RTT mouse model. (A) Elevated plus maze to evaluate anxiety-like behavior, the exposure to EE reestablished the preference for closed arms in Mecp2-/y mice. (B) Traveled distance in open field test, as a locomotion parameter, Mecp2-/y mice exposed to EE showed improved locomotion activity compared to Mecp2-/y mice exposed to SC. (C) Wire-hanging test to evaluate motor function, Mecp2-/y mice exposed to EE showed less failure in hanging from the wire in comparison with Mecp2-/y mice in SC. (D-G) Elevated dowel test to evaluate motor function, coordination and control, (D) Mecp2-/y mice in EE showed a similar number of falls as WT mice and fewer falls than Mecp2-/y mice in SC; EE exposure improved latency time (E) and the time of the first arrival (F) in Mecp2-/y mice, since they were similar to those exhibited by WT mice. (G) The number of arrivals of Mecp2-/y mice in SC were less than in WT mice, and this phenotype was completely prevented by the exposure to EE. (h) Motor coordination and learning was evaluated on the rotating cylinder of a rotarod, the poor motor coordination in Mecp2-/y mice were prevented by the exposure to EE, however motor learning was not recovered. These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p<0.05, ✱✱p <0.01, ✱✱✱p <0.001 and ✱✱✱✱p <0.0001.
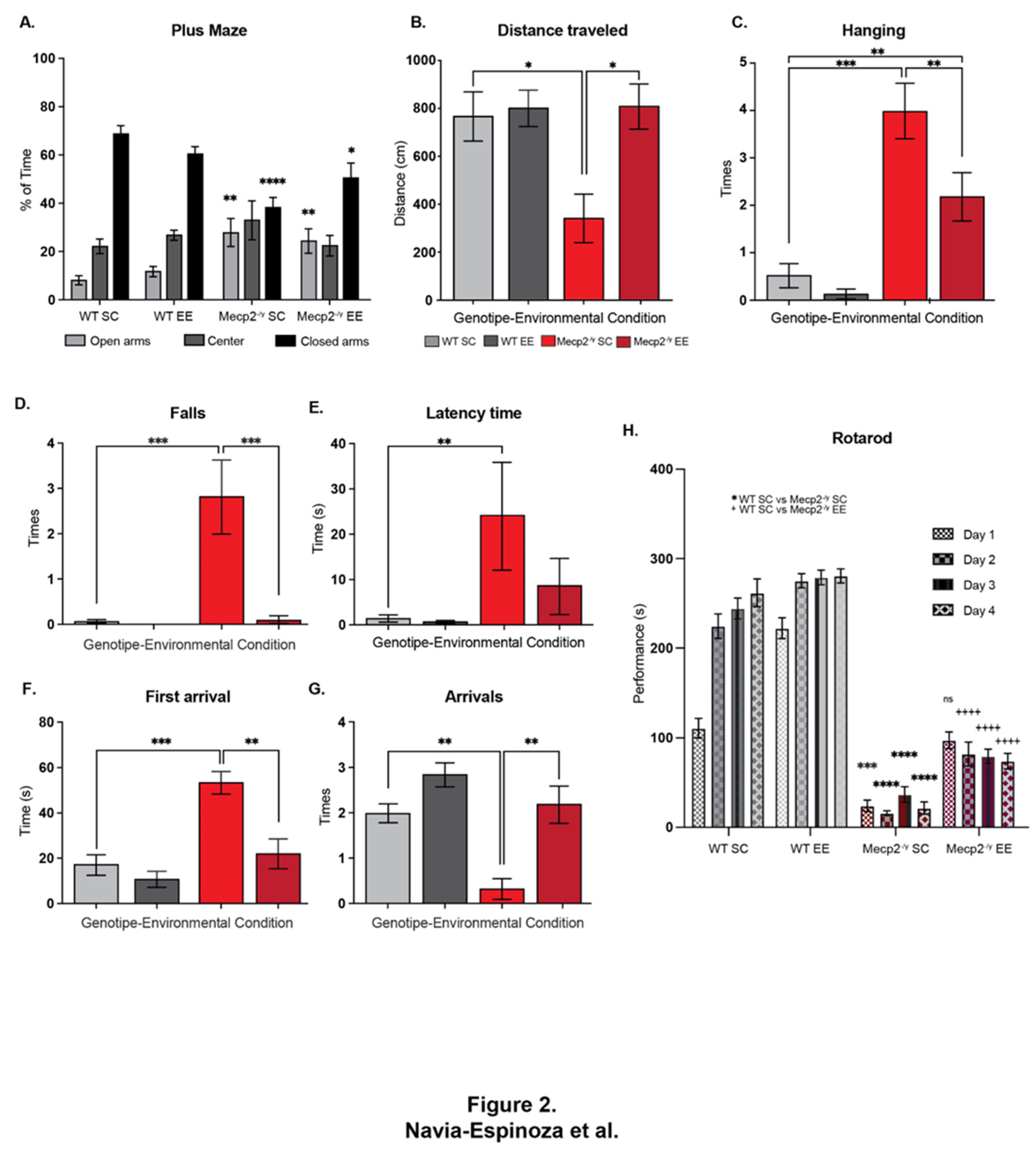
Figure 3.
Exposure to a neuronal plasticity-dependent paradigm decreases the synaptic deficit exhibited by an RTT mouse model. (A-D) M1-M2 motor cortex cytoarchitecture at 3 and 7 weeks of age of WT and Mecp2-/y mice exposed to either control or EE housing. (A) At 3 weeks of age, the motor cortex of Mecp2-/y mice is not far different from WT, (B-C) reduction of neuronal complexity and moderate astrogliosis in Mecp2-/y mice at 7 weeks of age which was improved by exposure to EE. (D-E) Evaluation of the dendritic length of pyramidal neurons of cortical layer 5, which was reduced in Mecp2-/y mice in SC in comparison with WT mice and this reduction was prevented by EE exposure. (F) The number of dendrites in pyramidal neurons was evaluated, Mecp2-/y mice exposed to EE had similar number that WT mice. (G-H) The dendritic spine density of second-order dendrites of pyramidal neurons from the motor cortex was evaluated, Mecp2-/y mice exhibit less spine density in comparison to that from WT in SC, which was prevented by EE exposure. These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱✱p <0.01, ✱✱✱p <0.001 and ✱✱✱✱p <0.001.
Figure 3.
Exposure to a neuronal plasticity-dependent paradigm decreases the synaptic deficit exhibited by an RTT mouse model. (A-D) M1-M2 motor cortex cytoarchitecture at 3 and 7 weeks of age of WT and Mecp2-/y mice exposed to either control or EE housing. (A) At 3 weeks of age, the motor cortex of Mecp2-/y mice is not far different from WT, (B-C) reduction of neuronal complexity and moderate astrogliosis in Mecp2-/y mice at 7 weeks of age which was improved by exposure to EE. (D-E) Evaluation of the dendritic length of pyramidal neurons of cortical layer 5, which was reduced in Mecp2-/y mice in SC in comparison with WT mice and this reduction was prevented by EE exposure. (F) The number of dendrites in pyramidal neurons was evaluated, Mecp2-/y mice exposed to EE had similar number that WT mice. (G-H) The dendritic spine density of second-order dendrites of pyramidal neurons from the motor cortex was evaluated, Mecp2-/y mice exhibit less spine density in comparison to that from WT in SC, which was prevented by EE exposure. These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱✱p <0.01, ✱✱✱p <0.001 and ✱✱✱✱p <0.001.
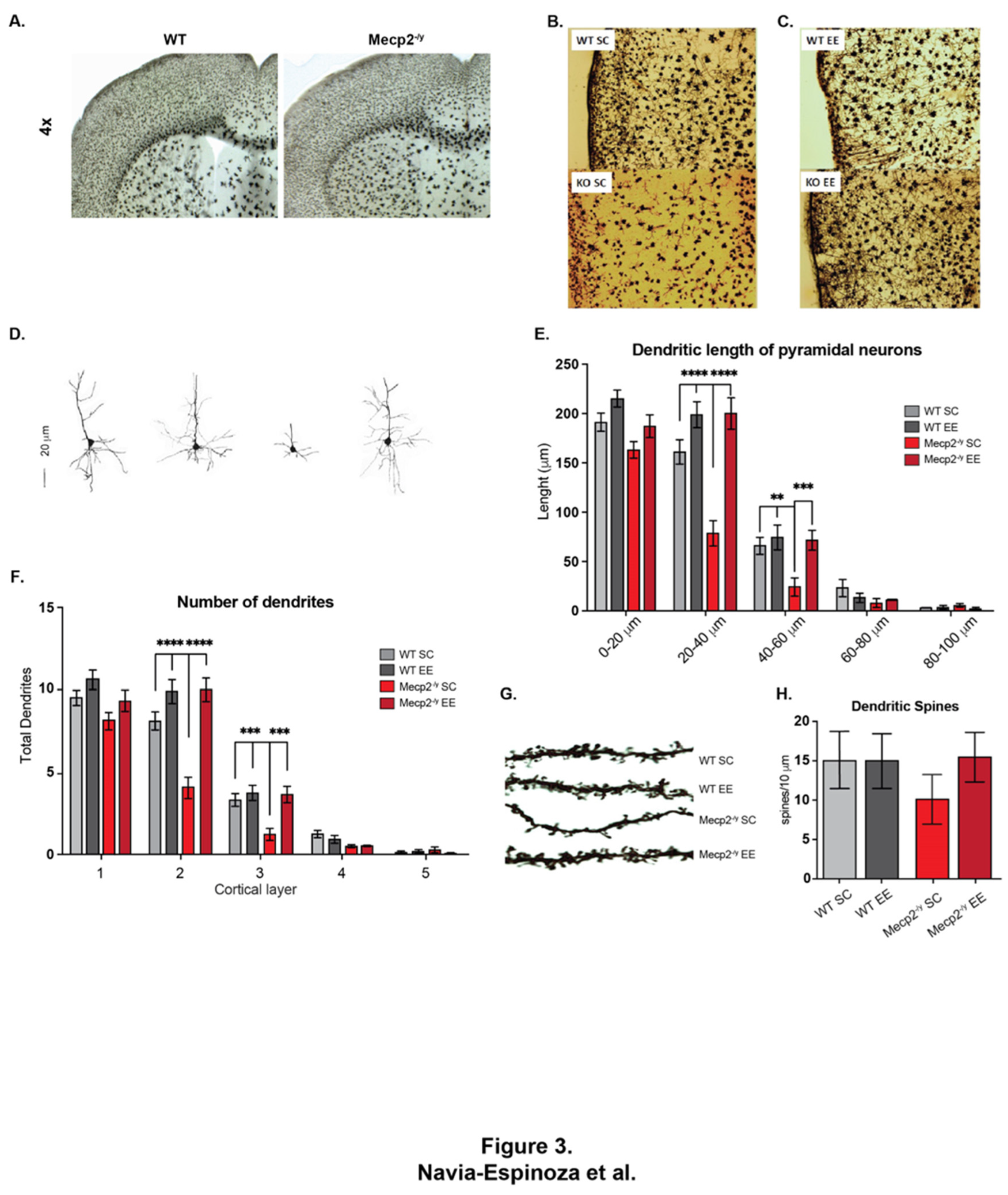
Figure 4.
Exposure to a neuronal plasticity-dependent paradigm can partially reestablish expression levels of genes related to cellular homeostasis. mRNA expression of GLAST (A) and GLT-1 (B) by RT-qPCR in WT mice exposed to SC, Mecp2-/y mice exposed to SC or EE, EE exposure failed to reestablish GLAST and GLT-1 expression levels in Mecp2-/y mice. (C-D) GLAST and GLT-1 protein levels are increased in Mecp2-/y mice exposed to SC compared to WT mice. (E-F) GLAST and GLT-1 are decreased in Mecp2-/y mice exposed to EE compared to those exposed to SC. (G) RT-qPCR of flip and flop splicing variants of Gria1, Gria2 and Gria3 genes, EE exposure restores the flip/flop ratio of Gria1 and Gria2 in Mecp2-/y mice. (H) mRNA expression of Irak1 by RT-qPCR, EE exposure reduced Irak1 expression in Mecp2-/y mice to levels like WT mice. These analyses were completed by (A, B, G, H) one-way ANOVA followed by Tukey’s multiple comparisons tests or (C-D) Mann-Whitney test. The levels of significance are shown by ✱p <0.05 and ✱✱✱p<0.001; ns, non-statistical differences.
Figure 4.
Exposure to a neuronal plasticity-dependent paradigm can partially reestablish expression levels of genes related to cellular homeostasis. mRNA expression of GLAST (A) and GLT-1 (B) by RT-qPCR in WT mice exposed to SC, Mecp2-/y mice exposed to SC or EE, EE exposure failed to reestablish GLAST and GLT-1 expression levels in Mecp2-/y mice. (C-D) GLAST and GLT-1 protein levels are increased in Mecp2-/y mice exposed to SC compared to WT mice. (E-F) GLAST and GLT-1 are decreased in Mecp2-/y mice exposed to EE compared to those exposed to SC. (G) RT-qPCR of flip and flop splicing variants of Gria1, Gria2 and Gria3 genes, EE exposure restores the flip/flop ratio of Gria1 and Gria2 in Mecp2-/y mice. (H) mRNA expression of Irak1 by RT-qPCR, EE exposure reduced Irak1 expression in Mecp2-/y mice to levels like WT mice. These analyses were completed by (A, B, G, H) one-way ANOVA followed by Tukey’s multiple comparisons tests or (C-D) Mann-Whitney test. The levels of significance are shown by ✱p <0.05 and ✱✱✱p<0.001; ns, non-statistical differences.
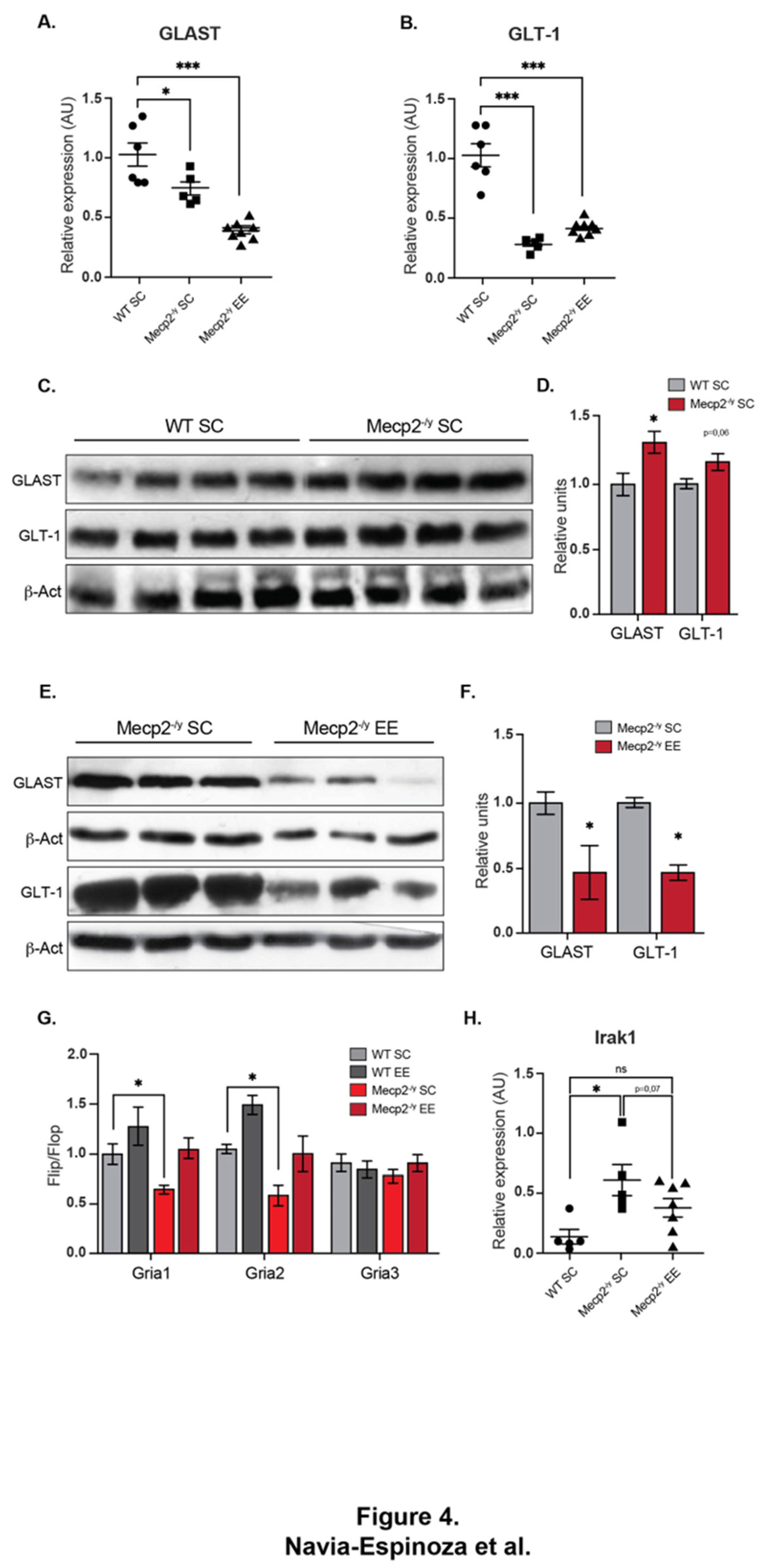
Figure 5.
Exposure to a neuronal plasticity-dependent paradigm ameliorates the alterations in neuromuscular junction distribution exhibited by an RTT mouse model. (A-B) Representative images of endplate distribution in the respiratory diaphragm muscle from WT and Mecp2-/y mice exposed to SC (A) or EE (B). (C) Mecp2-/y mice exposed to SC had increased average endplate width phenotype which was not observed in Mecp2-/y mice exposed to an enriched environment. (D) Histogram distribution of different endplate width ranges shows that the relative abundance of endplates from WT mice were not affected by the housing conditions; however, in Mecp2-/y mice exposed to EE the proportion of wider endplates (>800 mm width) was significantly reduced. These analyses were completed by (C) one-way ANOVA followed by Kruskal-Wallis’s multiple comparisons tests or (D) two-way ANOVA. The level of significance is shown by ✱p <0.05; ✱✱p<0.01.
Figure 5.
Exposure to a neuronal plasticity-dependent paradigm ameliorates the alterations in neuromuscular junction distribution exhibited by an RTT mouse model. (A-B) Representative images of endplate distribution in the respiratory diaphragm muscle from WT and Mecp2-/y mice exposed to SC (A) or EE (B). (C) Mecp2-/y mice exposed to SC had increased average endplate width phenotype which was not observed in Mecp2-/y mice exposed to an enriched environment. (D) Histogram distribution of different endplate width ranges shows that the relative abundance of endplates from WT mice were not affected by the housing conditions; however, in Mecp2-/y mice exposed to EE the proportion of wider endplates (>800 mm width) was significantly reduced. These analyses were completed by (C) one-way ANOVA followed by Kruskal-Wallis’s multiple comparisons tests or (D) two-way ANOVA. The level of significance is shown by ✱p <0.05; ✱✱p<0.01.
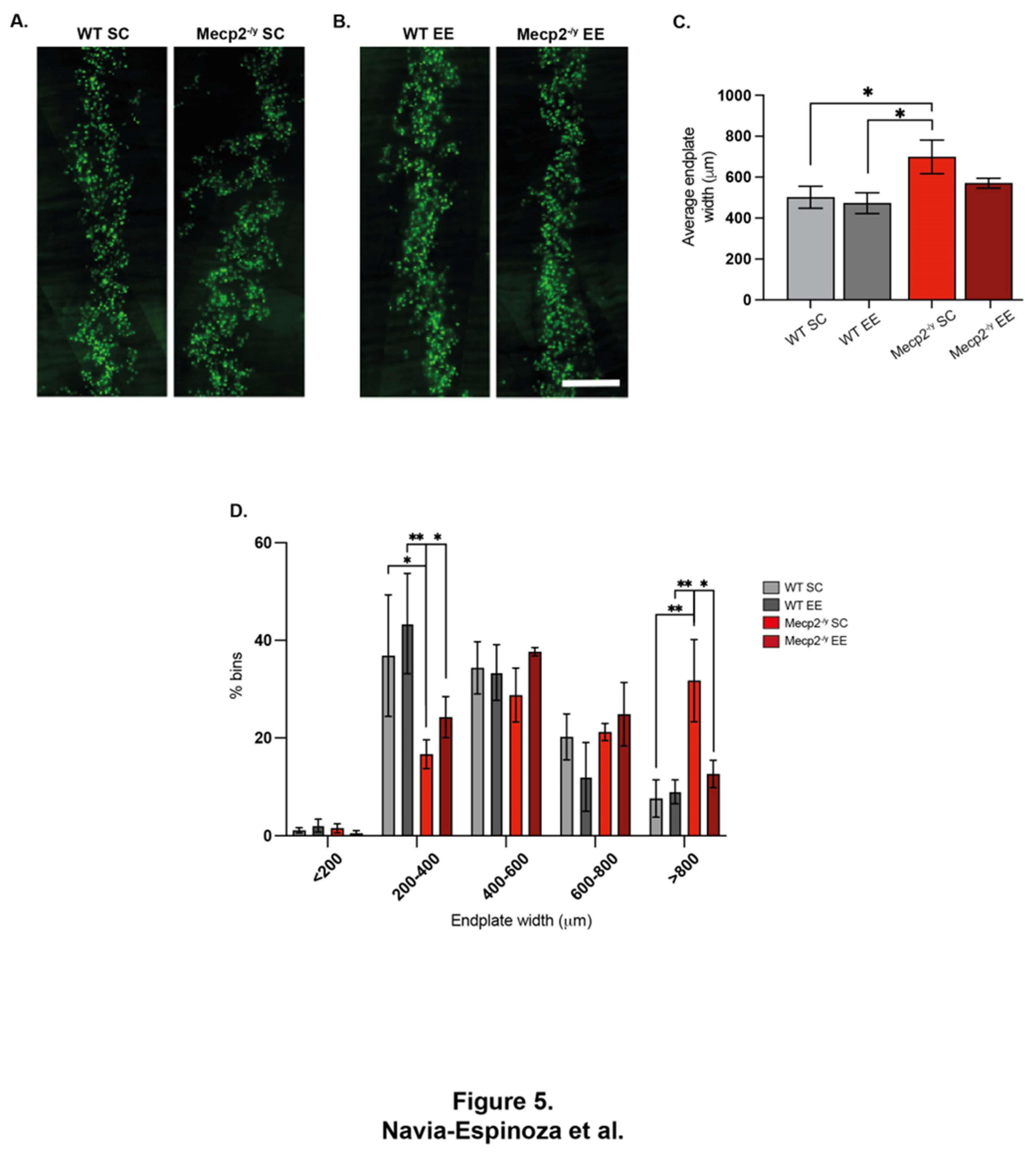
Figure 6.
The treatment with young plasma attenuates the RTT phenotype in a mouse model of the disease. (A) Lifespan of Mecp2-/y mice with or without IP plasma injection, IP plasma injection increases Mecp2-/y mice survival. (B-C) Phenotypic evaluation of Mecp2-/y mice, IP plasma injection attenuates the motor and neurological RTT-like symptoms progression evaluated in the (B) ledge test and (C) hindlimb clasping. (D) In the elevated plus maze to evaluate anxiety-like behavior, the IP plasma treatment did not prevent Mecp2-/y mice preference for the open arm of the maze. (E-F) In the elevated dowel test to evaluate motor coordination, IP plasma treatment prevented the increased time of the first arrival (E) and the increased number of falls (F) in Mecp2-/y mice. These analyses were completed by (A) Simple survival analysis Mantel-Cox test followed by Wilcoxon test, and (B-F) two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p <0.05 ✱✱p or ++p <0.01, ✱✱✱p or ###p <0.001 and ✱✱✱✱p or ++++p <0.001; ns, non-statistical differences.
Figure 6.
The treatment with young plasma attenuates the RTT phenotype in a mouse model of the disease. (A) Lifespan of Mecp2-/y mice with or without IP plasma injection, IP plasma injection increases Mecp2-/y mice survival. (B-C) Phenotypic evaluation of Mecp2-/y mice, IP plasma injection attenuates the motor and neurological RTT-like symptoms progression evaluated in the (B) ledge test and (C) hindlimb clasping. (D) In the elevated plus maze to evaluate anxiety-like behavior, the IP plasma treatment did not prevent Mecp2-/y mice preference for the open arm of the maze. (E-F) In the elevated dowel test to evaluate motor coordination, IP plasma treatment prevented the increased time of the first arrival (E) and the increased number of falls (F) in Mecp2-/y mice. These analyses were completed by (A) Simple survival analysis Mantel-Cox test followed by Wilcoxon test, and (B-F) two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p <0.05 ✱✱p or ++p <0.01, ✱✱✱p or ###p <0.001 and ✱✱✱✱p or ++++p <0.001; ns, non-statistical differences.
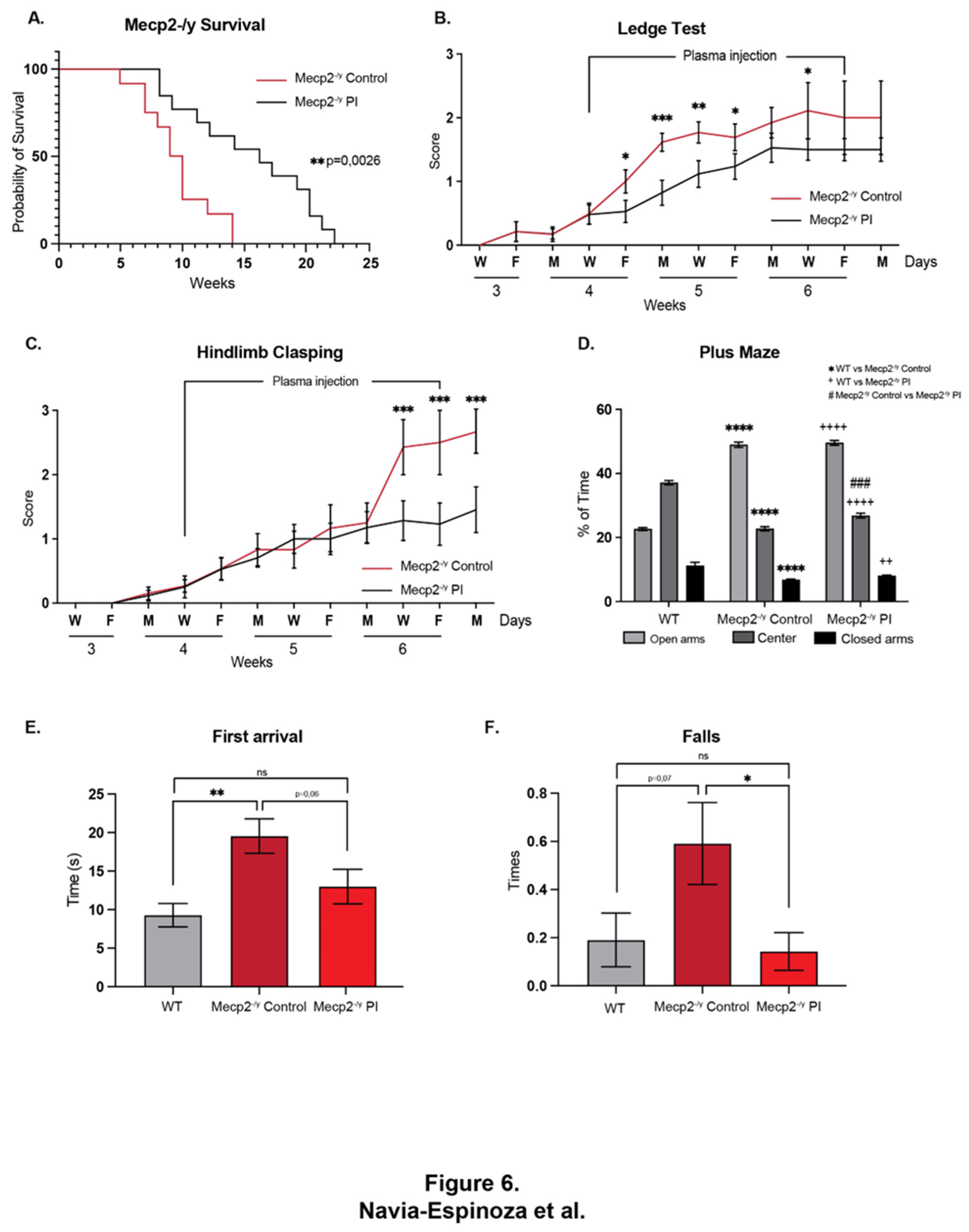
Figure 7.
The treatment with young plasma decreases the synaptic deficit exhibited by an RTT mouse model. (A) Golgi staining of Mecp2-/y control and IP plasma injection treated mice and their WT littermates. Plasma treatment prevented the diminished brain architecture observed in Mecp2-/y mice in the control group. IP plasma treatment almost reestablished the thickness of the corpus callosum (B) and motor cortex (C) to like that observed in the WT littermates. (D) Moderate signs of astrogliosis observed in Mecp2-/y mice, which was attenuated by IP plasma injection. (E) Neuronal cytoarchitecture of pyramidal neurons was reestablished, as evaluated by dendritic length (F) and dendritic arborization complexity (G). These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p or +p <0.05, ✱✱p, ++p or ##p <0.01, ✱✱✱p or +++p <0.001 and ✱✱✱✱p, ++++p or ####p<0.001.
Figure 7.
The treatment with young plasma decreases the synaptic deficit exhibited by an RTT mouse model. (A) Golgi staining of Mecp2-/y control and IP plasma injection treated mice and their WT littermates. Plasma treatment prevented the diminished brain architecture observed in Mecp2-/y mice in the control group. IP plasma treatment almost reestablished the thickness of the corpus callosum (B) and motor cortex (C) to like that observed in the WT littermates. (D) Moderate signs of astrogliosis observed in Mecp2-/y mice, which was attenuated by IP plasma injection. (E) Neuronal cytoarchitecture of pyramidal neurons was reestablished, as evaluated by dendritic length (F) and dendritic arborization complexity (G). These analyses were completed by two-way ANOVA followed by Tukey’s multiple comparisons tests. The levels of significance are shown by ✱p or +p <0.05, ✱✱p, ++p or ##p <0.01, ✱✱✱p or +++p <0.001 and ✱✱✱✱p, ++++p or ####p<0.001.
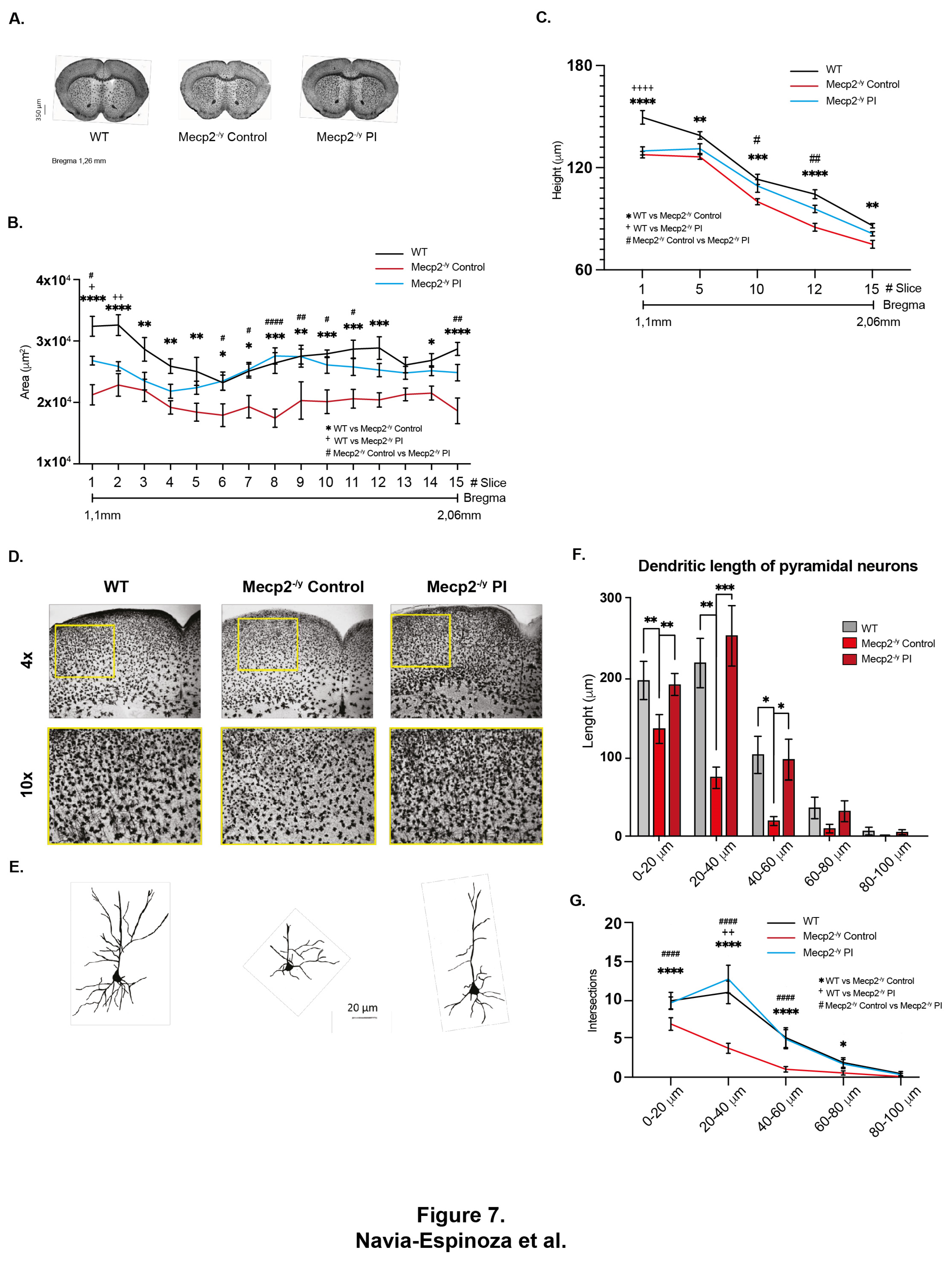

Table 1.
Primers used for RT-qPCR.
| Target name | Forward Primer | Reverse Primer |
|---|---|---|
| GLAST | ACGGTCACTGCTGTCATT | TGTGACGAGACTGGAGATGA |
| GLT-1 | CTGGTGCAAGCCTGTTTCC | GCCTGTTCACCCATCTTCC |
| Irak1 | ACTACATATGCTGTGAAGAGA | CTCATCCAGAAGCACGTTAGA |
| Gria1 flip | ACACCATGAAAGTGGGAGGTAACT | ACTGGTCTTGTCCTTACTTCCGGA |
| Gria1 flop | GTCCGCCCTGAGAAATCCA | GCACTCGCCCTTGTCGTA |
| Gria2 flip | ACACCATGAAAGTGGGCGGCAACC | ACTGGTCTTTTCCTTACTTCCCGA |
| Gria2 flop | ACACCATGAAAGTGGGCGGCAACC | ACTGGTCTTTTCCTTGGAATCACC |
| Gria3 flip | ATACGATGAAAGTTGGTGGAAATC | ACTGGTCTTGTCCTTACTCCCGGA |
| Gria3 flop | ATACGATGAAAGTTGGTGGAAATC | ACTGGTCTTGTCCTTGGAGTCACC |
| Cyc | GGCAATGCTGGACCAAACACAA | GTAAAATGCCCGCAAGTCAAAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



