Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Oxytocin, known as the “love hormone,” is produced in the posterior hypothalamus and participates in numerous physiological processes, including social intelligence, sexual activity, and metabolism. SARS-CoV-2 virus, the etiologic agent of the COVID-19 pandemic, has been shown to suppress oxytocin release from the intestinal epithelial cells, resulting in microbial migration outside of the gut lumen.
Limosilactobacillus reuterii, a commensal known for producing oxytocin, normalizes the gut barrier function and exerts antiviral properties, including against the SARS-CoV-2 virus. This may explain the beneficial effect of oxytocin in schizophrenia, a condition associated with a dysfunctional gut barrier and microbial translocation from the gastrointestinal tract into the systemic circulation.
The molecular underpinnings of oxytocin in the central nervous system are not entirely clear. However, as viruses target tight junctions, the molecular Velcro that holds the barrier cells together, oxytocin enhances the barrier function.
In this mini-view, we summarize what is known about the role of oxytocin in schizophrenia and discuss natural and synthetic compounds that could optimize gut barrier permeability.
Keywords:
schizophrenia
; microbiome
; endocrine organ
; virus
; autoantibodies
Introduction
The COVID-19 pandemic was an eye-opener for many medical disciplines. It highlighted viral exploitation of the physiological cellular ingress routes (endocytosis), generally utilized for signaling, receptor recycling, engulfment of xenobiotics, and cell-cell adhesion. These entry pathways play an essential role in cancer, neurodegenerative disorders, and schizophrenia (SCZ).
Viruses like SARS-CoV-2 exploit other cellular products, including hormones and neurotransmitters. For example, the Japanese Encephalitis Virus (JEV) usurps dopamine (DA) receptors to enter cells, therefore disrupting dopaminergic signaling [1]. Moreover, viruses can hijack cellular senescence, an antitumor program in which the cell permanently exits the cell cycle to deter malignant transformation.
Due to its antiviral properties, oxytocin (OXT) is targeted by the SARS-CoV-2 virus, resulting in decreased signaling with OXYTOCIN receptors (OXRs), which affects among other things, male and female fertility [2,3,4]. Indeed, patients with SCZ have fewer offspring than the general population, which may, at least partially, result from dysfunctional OXYTOCIN signaling [5]. This may be further substantiated by the COVID-19-related drop in birth rate in many countries, including the US [6,7].
The connection between viruses and SCZ has been documented for several decades. For example, women pregnant during the US rubella epidemic in 1964 gave birth to offspring that developed autism spectrum disorders (ASDs) or SCZ more often than the population at large, suggesting that many viruses, probably including COVID-19, may contribute to these sequelae [8]. Other studies found that dormant viruses, such as human endogenous retroviruses (HERVs), could promote SCZ [9]. Moreover, molecular mimicry between the M2 protein of influenza A virus and N-methyl-D-aspartate receptors (NMDARs) can explain the “autoantibodies” against this protein found in some SCZ patients, further linking viruses to chronic psychosis [10,11]. These “autoantibodies” are likely conventional immunoglobulins directed at the M2 viral protein. Moreover, some gut microbes, including Helicobacter pylori, express NMDA receptors, further enhancing the fact that translocated microbes can elicit the formation of antibodies against these proteins.
The gut microbes are known to produce all the neurotransmitters and hormones the host generates, so the microbiome is often conceptualized as an endocrine organ [12,13]. Limosilactobacillus reuteri (L. reuteri), one of the OXYTOCIN-producing gut microbes, has been less abundant in humans over the past two decades, probably accounting for the increased prevalence of autoimmune disorders [14]. L. reuteri is found in the GI tract, urinary tract, skin, and breast milk, where it exerts anti-inflammatory, antimicrobial, and antianxiety actions and produces vitamin B12 and histamine, contributing to the GI tract's homeostasis [15].
OXYTOCIN is a peptide hormone synthesized in the hypothalamic paraventricular (PVN) and supraoptic (SON) nuclei and stored in the posterior pituitary. Aside from L. reuteri, intestinal epithelial cells (IECs) produce OXYTOCIN, especially when exposed to the GI tract hormone secretin [16]. This may explain the documented beneficial effect of secretin in refractory SCZ [17]. OXYTOCIN signals with OXRs spread throughout the brain and are more numerous in the hypothalamus [18].
To function adequately, OXRs require cholesterol, a lipid that delays receptor degradation. Thus, OXYTOCIN signaling is likely deficient at low cholesterol levels [19]. In addition, statins can exhibit extracerebral adverse effects, such as infertility and muscle breakdown [20,21]. Moreover, increasing evidence suggests that the beneficial effects of statins on endothelial cells may be independent of their cholesterol-lowering properties [22].
The prosocial role of OXYTOCIN and the fact that abundant OXYTOCIN receptors are expressed in the insular cortex (IC) and anterior cingulate cortex (ACC) has led many researchers and clinicians to see a functional similarity between OXYTOCIN and von Economo neurons (VENs), the large bipolar cells that project to a distant area of the brain [23]. Indeed, VENs are part of the salience network (SN), a neuronal assembly that shifts attention from exteroception to interoception or from the central executive network (CEN) to the default mode network (DMN), depending on the stimulus relevance [24].
Although not officially included among senolytic or xenomorphic agents, OXYTOCIN meets the criteria for the latter. Senotherapeutics are divided into senolytic compounds that accelerate the elimination of senescent cells and xenomorphics that delete the senescence markers, “rejuvenating” the cell.
This mini-view looks closely at potential interventions based on the microbial translocation hypothesis.
Cellular Senescence
Under physiological circumstances, after replicating 40-60 times, somatic cells undergo senescence, marked by proliferative arrest and a rewired metabolism. Exceptions to this rule are cancer cells and probably stem cells, which can divide indefinitely. For example, HeLa cells, derived from a 1950s cancer patient, are still thriving in labs worldwide and have been used recently for research on the SARS-CoV-2 vaccine [25]. Aside from replicative senescence, cells can activate the senescence program when in danger of malignant transformation or damaged DNA needs repair.
Cellular senescence is marked by irreversible replication arrest, resistance to apoptosis, active metabolism, and the release of a toxic secretome known as the senescence-associated secretory phenotype (SASP). The accumulation of senescent cells is believed to contribute to organismal aging, as SASP can spread the senescent phenotype locally and systemically. For example, aging endothelial cells (ECs) release SASP directly into the systemic circulation, disseminating senescence throughout the body [26].
Several studies have shown that although senescence itself protects against malignant transformation, SASP maintains a degree of low-grade inflammation that may predispose to cancer.
Phagocytes clear senescent cells in a process known as efferocytosis. When efferocytosis is defective, senescent cells accumulate, triggering inflammation [27]. In patients with severe mental illness, efferocytosis is defective, leading to premature aging and low-grade inflammatory responses. Indeed, SCZ patients live on average 15-20 years shorter than the general population and develop age-related illnesses earlier in life, leading some researchers to conceptualize SCZ as a “segmental progeria” [28]. Along this line, telomeres are shorter in patients with SCZ, and senescent markers, such as p21, p16, and SA-β-galactosidase activity, are frequently elevated.
OXYTOCIN opposes cellular senescence, especially the SASP, and isolation-induced premature aging observed during the pandemic lockdown [29]. Along this line, intranasal OXYTOCIN has been shown to improve social interaction and communication in children with ASD, highlighting a novel therapeutical strategy for this condition. Aside from eliciting pro-social behavior, OXYTOCIN exerts direct antiviral effects, especially against the SARS-CoV-2 virus, as reported by several studies [30]. Indeed, this action brought OXYTOCIN into the spotlight of research during the COVID-19 pandemic.
In the GI tract, OXYTOCIN receptors are highly represented in the myenteric plexus and are involved in motility, inflammation, and barrier permeability. For example, OXYTOCIN receptors prevent microbial translocation outside of the GI tract, an action canceled by vagotomy, a finding that implicates cholinergic neurotransmission, probably via signal transducer and activator of transcription 3 (STAT3), in OXYTOCIN release [31,32]. In addition, as OXYTOCIN upregulates IL-10, an anti-inflammatory cytokine that shares its receptor with IL-22, it likely upregulates this “guardian of the gut barrier,” preventing translocation [33]. Moreover, like IL-22, OXYTOCIN mediates wound healing, suggesting that it also improves biological barriers.
Schizophrenia Outcome Studies
Gray matter volume (GMV) reduction, a hallmark of SCZ demonstrated by many neuroimaging studies, is seldom considered in the etiopathogenesis of this disorder, probably because it is refractory to most of the available treatments. GMV loss is in line with SCZ outcome studies, which show that only a minority of patients reach sustained recovery, suggesting that brain volume atrophy may progress unhindered by the antipsychotic drugs we currently utilize [34]. White matter is also depleted in SCZ, although less than the gray matter, and likely reflects the damaged myelin.
GMV reduction in SCZ, associated with aggression and violence, occurs in both medicated and unmedicated SCZ patients, suggesting that aggressive behavior is an inherent part of the disease and not caused by the antipsychotic treatment only [35,36,37].
OXYTOCIN treatment may improve SCZ outcomes, as it was shown to preserve gray matter and ameliorate traumatic brain injury (TBI) and epilepsy [38,39]. This may alter the disease course, improving functional recovery.
With current SCZ treatments, 33% of patients relapse during the first 12 months after an initial psychotic episode, 26% remain homeless at two-year follow-up, while five years after the first psychotic outbreak, only 10% are employed [40,41,42].
Clinically, the natural progression of SCZ evolves in four stages: an asymptomatic phase followed by a prodrome with mild but not overly psychotic symptoms. Phase III is manifested as frank psychosis and comprises the first psychotic episode and subsequent relapses. In stage IV, usually after midlife, the positive symptoms gradually subside and are replaced by negative and cognitive manifestations [43].
The ”Love Hormone” and The Fat
In humans, cholesterol comprises 35% of cell membranes and is found in the cell membrane’s lipid bilayer, dispersed among phospholipids. Cholesterol provides stability to cells and organelles by maintaining an optimal balance between membrane fluidity and rigidity (Figure 1). In neurons, cholesterol is a significant driver of dendritic spine and neurite growth, implicating this lipid in memory and cognition [44]. In this regard, loss of dendritic arborization in inhibitory interneurons was associated with aggression in patients with SCZ, suggesting that this disorder may be caused by a lipidome dysfunction [45].
Cholesterol is an essential allosteric modulator of OXYTOCIN receptors (OTRs), stabilizing this high-affinity bond. This raises the possibility of statins altering OXYTOCIN signaling, precipitating iatrogenic aggression. Indeed, several studies reported statin-induced violence, suggesting that plasmalogen, the “natural statin,” should be used for the management of hypercholesterolemia in psychiatric patients [46,47].
In a previous study, we hypothesized that low cholesterol and hypovitaminosis D contribute to GMV reduction, a hallmark of SCZ and aggression. We hypothesized further that a subgroup of patients with increased expression of the ATP binding cassette subfamily A member 1 (ABCA1) gene is more vulnerable to cholesterol-lowering strategies [48]. Hypovitaminosis D was demonstrated in over 95% of forensic patients with SCZ, implicating this nutrient in neuropathology [49]. Low levels of cholesterol, a vitamin D precursor, were associated with both violent behavior and suicide, highlighting further the role of lipidome in this pathology [50,51,52].
The brain is the cholesterol-richest organ in the body. In the central nervous system (CNS), cholesterol is a component of the lipid bilayer of neuronal membranes and the myelin sheath. Aggressive and violent behaviors in patients with SCZ are directly correlated with gray matter loss, especially in the right superior frontal cortex, left inferior parietal region, insular cortex (IC), and ventromedial prefrontal cortex (VMPFC)[53,54,55]. In contrast, suicidal behavior (self-aggression) was associated with white matter loss, implicating defective remyelination in this pathology [56].
Cholesterol depletion was previously demonstrated to disrupt glutamate α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors, leading to the loss of synapses and dendritic spines, pathologies documented in SCZ variants marked by violent behaviors [57,58].
Several studies have found that OXYTOCIN can trigger reactive anger and aggression, especially in SCZ patients [59,60,61]. Indeed, despite promoting prosocial behavior, OXYTOCIN may, in some situations, enhance aggression. However, the relationship with low cholesterol or lipid peroxidation requires more studies. Along this line, several articles have associated SCZ with lipid peroxidation in cells, including the neurons. At the same time, lipophilic antipsychotic drugs, such as phenothiazines, infiltrate the membrane lipid bilayer, repairing the lipids with their antioxidant properties [62,63]. Several new phenothiazines oppose lipid peroxidation, highlighting the dopamine-independent antipsychotic action of these drugs [64]. Indeed, novel phenothiazine derivatives with potent antioxidant properties are available to treat heart disease and cancer. To our knowledge, despite the phenothiazine core, these agents were never evaluated for mental illness. For example, tetracyclic and pentacyclic phenothiazine derivatives are potent antioxidants and free radical scavengers capable of repairing the membrane lipids and rescuing cells, including the neurons [65]. Phenazines are natural phenothiazines generated by various species of soil and marine microbes and exhibit antibiotic, anticancer, and antipsychotic properties [66].
Aryl Hydrocarbon Receptor, the New Kid on the Block
The aryl hydrocarbon receptor (AhR) was initially known as the dioxin receptor. However, many ligands at this protein have been identified over the years, some of which are relevant to neuropathology. For example, serotonin (5-HT), melatonin, and dopamine (DA) are upstream AhR ligands, bringing this protein into the psychiatric arena. In addition, AhR is the master regulator of cellular senescence because it can sense endogenous and exogenous metabolites, xenobiotics, and toxicants, inducing premature molecular aging to lower the risk of malignant transformation.
AhR negatively regulates lactate and its posttranslational modification, lactylation. Consequently, dysfunctional AhR may drive both premature neuronal and glial aging, as well as the excessive lactylation documented in SCZ and other psychotic disorders [67]. For example, senescent microglia with excessive histone 3 (H3K18) lactylation were demonstrated to adopt a neurotoxic phenotype, eliminating healthy synapses and neurons, as documented in SCZ and AD [68].
Translocated microbes trigger iron sequestration in host macrophages, a phenomenon known as nutritional immunity. Nutritional immunity aims to withhold iron from the invading pathogens, decreasing infectivity [69]. However, low circulatory iron often causes bacteria in host tissues, including the brain, to adopt a dormant phenotype [70]. For example, dormant brain microbes were documented in Alzheimer’s disease (AD), while a virome (viral microbiome) was recently identified in the SCZ brains (Broadman area 46) [71,72]. The dormant microbes can be reactivated when iron becomes available, causing neuropathology. Moreover, iron regulatory proteins, such as hepcidin, are AhR ligands, emphasizing a previously unknown relationship between AhR and nutritional immunity [73].
AhR is located at the biological barriers and comes readily into contact with endogenous and exogenous nutrients and toxins that bind this receptor with various affinities, eliciting different degrees of activation. Excessive AhR activation likely disrupts the TJ molecules, opening paracellular pathways used by microbes or their components to migrate outside the lamina propria [74]. In this regard, SCZ with negative symptoms was associated with antibodies against translocated Hafnei alvei, Pseudomonas aeruginosa, Pseudomonas putida, and Klebsiella pneumonia, which are proof of concept of microbial migration in SCZ.
Viruses also induce cellular senescence by manipulating AhR through their antigens. For example, the SARS-CoV-2 virus usurps OXYTOCIN via OXYTOCIN-activating motifs such as calcium calmodulin kinase II (CaMKII). At the molecular level, OXYTOCIN accumulates and phosphorylates CaMKII, lowering the circulatory levels of this hormone. Moreover, the S antigen of the SARS-CoV-2 virus contains soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptors (SNARE) repeats that can interfere with the vesicular release of OXYTOCIN, further lowering the systemic level of this hormone [75] (Figure 3).
Molecular imitation of placental proteins by viral arginine repeats, HAP2, tubulin, tau, and human endogenous retroviruses (HERVs) facilitates both the infection and the development of antibodies against host proteins.
Taken together, molecular mimicry of reproductive proteins leads to reduced birth rates as documented worldwide. Viral repeats (motifs) are known AhR activators, an action that manipulates this protein into a pro-viral role. Active AhR lowers interferon-gamma, a potent antiviral defense [76,77]. Therefore, AhR inhibitors exert not only anti-translocation but also antiviral properties.
Potential Interventions
The microbial translocation hypothesis opens new therapeutic strategies beyond dopamine and the synapse. These strategies include senotherapeutics, barrier enhancers, and AhR inhibitors.
Senotherapeutics
SCZ has been associated with premature molecular aging, which is reflected in decreased longevity by 15-20 years compared to the general population. Premature molecular aging likely results in dysfunctional TJs and subsequent microbial translocation in the host’s tissues and organs.
Senotherapeutic drugs are comprised of senolytic agents (which promote the elimination of senescent cells) and senomorphic agents (which delete senescent markers). Senotherapeutics are underutilized in SCZ, but we believe they could be beneficial. For example, eliminating neurotoxic microglia could avert neurodegeneration and SCZ by sparing neuronal and glial cells.

Table 1.
Common natural and synthetic senolytic and senomorphic drugs.
| Senolytic drugs | Source | Mechanism | References |
|---|---|---|---|
| Kaempferol | Fruits and vegetables | GSK-3 beta inhibitor | [78] |
| Berberine | Oregon grape, phellodendron, and tree turmeric | Ceramide inhibitor | [79]. |
| Lycopene | Grape skin, guava, grapefruit, blueberries, and tomatoes | Scavenging of reactive oxygen species, enhancement of detoxification systems | [80]. |
| Fisetin | Strawberries, onions, apples, mangoes, persimmons, and kiwis | inhibiting the activity of nuclear factor-kappa B (NF-κB) and MAPK. | [81] |
| Senomorphic drugs | |||
| Rapamycin | bacterium Streptomyces hygroscopicus | mTOR inhibition | [82]. |
| Acarbose | Soil bacteria | PPARγ upregulation | [83]. |
| SIRT-1 | Grapes, berries, apples, and other fruits | Lowers oxidative stress | [84] |
| Fluvastatin and Valsartan | Synthetic | Increase telomerase activity | [85]. |
| KU-60019 | Synthetic | Improves mitochondrial function | [86]. |
In the past, it was believed that cellular senescence was irreversible. However, newer studies have found that inhibiting 3-phosphoinositide-dependent protein kinase-1 (PDK1) can reverse this phenotype [87]. For this reason, PDK1 inhibitors will likely play a significant role in SCZ.
Barrier Enhancers
Premature cellular senescence in SCZ affects most cell types, including those comprising the biological barriers. The senescent phenotype disrupts the TJs, enabling the translocation of gut bacteria into the host circulation. The GI tract barrier is comprised of epithelial and endothelial cells. IECs produce OXYTOCIN when prompted by secretin, a GI hormone the enterochromaffin cells produce. Earlier studies have associated secretin with symptomatic improvement of refractory SCZ, suggesting an indirect action via OXYTOCIN [88]. Along this line, a recent study found that gut microbes influence OXYTOCIN release, implicating commensal flora in the biosynthesis of this hormone [89].
Indeed, it has been established that human commensal Limosilactobacillus reuterii produces OXYTOCIN, protecting the gut barrier [90]. For this reason, probiotics with L. reuterii should be supplemented for SCZ patients. In addition, previous studies found that Bifidobacterium breve A-1 ameliorated depression, anxiety, and cognition, combined with L. reuterii, could benefit SCZ. In addition, both microbes likely upregulate IL-22, “the guardian of the gut barrier,” further averting microbial translocation [91].
During the HIV epidemic in the 1980s, IL-22 deficit was linked to increased intestinal permeability and HIV-induced massive translocation of gut microbes into host tissues. This occurred as the virus targeted innate lymphoid cells type 3 (ILC-3), producers of IL-22. Depletion of this cytokine led to microbial migration outside the gut lumen. For this reason, we proposed human recombinant IL-22 as a novel SCZ treatment [92]. Other interventions for enhancing the gut barrier include serine proteases, such as nafamostat mesylate and camostat mesylate, known for decreasing paracellular spaces' size [93]. Indigo and indirubin, Chinese herbal medicines, are enhancers of the gut barrier that are beneficial in IBD and probably SCZ.
OSU 03012 and Other PDK1 Inhibitors
Membranes separate body compartments, maintaining gradients and electrical charges that drive cellular machinery, ultimately making life possible. Plasma and organelle membranes comprise a lipid bilayer, and lipid peroxidation can alter the biophysical properties of membranes, disrupting the surface receptors and neurotransmission.
Due to their ability to reverse cellular senescence, PDK1 inhibitors are of extreme interest in SCZ, a disorder marked by premature aging and early mortality [94,95]. Indeed, several PDK1 inhibitors, including alpha-lipoic acid N acetylcysteine (NAC), have been found to lower lipid peroxidation and are currently being evaluated as SCZ treatments [96,97,98]( NCT03788759).
Akt kinase is activated in cell membranes by PI3K, which stimulates PDK1, a master kinase recruited to the plasma membrane via its C-terminal pleckstrin homology (PH) domain. Interestingly, PH polymorphisms were previously associated with SCZ, further implicating PDK1 in this disorder [99].
The aging brain retains iron, which can enhance the oxidation of membrane lipid bilayer. This pathology could be ameliorated or reversed by MLR [101,102,103]. Interestingly, aside from signaling with its receptors, DA also binds directly to membrane lipids, triggering peroxidation via 6-hydroxydopamine (6-OHDA), an established neurotoxin [104,105].
We propose replacing oxidized membrane lipids with OSU 03012, a synthetic PDK1 inhibitor, and natural exogenous glycerophospholipids. We discussed MLR in other articles and will not repeat it here (Sfera A)( Del Campo CMZ). Initially described by Professor Garth Nicolson, who studied Gulf War Syndrome since 1990s, MLR was expanded to fatiguing disorders. We saw potential benefits of MLR in SCZ, a condition marked by premature cellular senescence and extensive oxidation of membrane lipids.
OSU-03012 is a celecoxib derivative that exerts antiproliferative, antibacterial, and antiviral properties and shares many characteristics of antipsychotic drugs (Table 2).
Devoid any COX-inhibiting activity, OSU-03012 binds to and inhibits PDK1 (Figure 4) [115]. Subsequently, the phosphorylation and activation of Akt are inhibited, disrupting the PI3K/Akt signaling pathway, a marker of tumorigenesis and SCZ.
Conclusions
Synthesized in the CNS and the gut, OXYTOCIN is far from only being the “hormone of love” and prosocial behaviors. In the GI tract, OXYTOCIN maintains the homeostasis of the gut barrier, preventing microbial translocation and likely improving the outcome of SCZ.
Interventions to upregulate OXYTOCIN, including senotherapeutics, AhR, and PDK-1 inhibitors, represent new strategies that could be helpful in SCZ and IBD. A particular intervention, MLR with OSU-03012 and human recombinant IL-22, may optimize the gut and BBB, eliminating the dormant CNS microbes.
References
- Simanjuntak Y, Liang JJ, Lee YL, Lin YL. Japanese Encephalitis Virus Exploits Dopamine D2 Receptor-phospholipase C to Target Dopaminergic Human Neuronal Cells. Front Microbiol. 2017 Apr 11;8:651. [CrossRef]
- Lu X, Liu Y, Zhang J, Wu X, Li X. Oxytocin increases pregnancy rates after fixed-time artificial insemination in Kazak ewes. Reprod Domest Anim. 2021 Jun;56(6):942-947. [CrossRef]
- KNIGHT, T. W. (1974). THE EFFECT OF OXYTOCIN AND ADRENALINE ON THE SEMEN OUTPUT OF RAMS. Reproduction, 39(2), 329-336. Retrieved Nov 12, 2024, from . [CrossRef]
- Wang SC, Zhang F, Zhu H, Yang H, Liu Y, Wang P, Parpura V, Wang YF. Potential of Endogenous Oxytocin in Endocrine Treatment and Prevention of COVID-19. Front Endocrinol (Lausanne). 2022 May 3;13:799521. [CrossRef]
- Stevenson JR, McMahon EK, Boner W, Haussmann MF. Oxytocin administration prevents cellular aging caused by social isolation. Psychoneuroendocrinology. 2019 May;103:52-60. [CrossRef]
- Winkler-Dworak M, Zeman K, Sobotka T. Birth rate decline in the later phase of the COVID-19 pandemic: The role of policy interventions, vaccination programmes, and economic uncertainty. Hum Reprod Open. 2024 Sep 10;2024(3):hoae052. [CrossRef]
- Ullah MA, Moin AT, Araf Y, Bhuiyan AR, Griffiths MD, Gozal D. Potential Effects of the COVID-19 Pandemic on Future Birth Rate. Front Public Health. 2020 Dec 10;8:578438. [CrossRef]
- Shuid AN, Jayusman PA, Shuid N, Ismail J, Kamal Nor N, Mohamed IN. Association between Viral Infections and Risk of Autistic Disorder: An Overview. Int J Environ Res Public Health. 2021 Mar 10;18(6):2817. [CrossRef]
- Leboyer M, Tamouza R, Charron D, Faucard R, Perron H. Human endogenous retrovirus type W (HERV-W) in schizophrenia: A new avenue of research at the gene-environment interface. World J Biol Psychiatry. 2013 Mar;14(2):80-90. [CrossRef]
- Kolocouris A, Spearpoint P, Martin SR, Hay AJ, López-Querol M, Sureda FX, Padalko E, Neyts J, De Clercq E. Comparisons of the influenza virus A M2 channel binding affinities, anti-influenza virus potencies and NMDA antagonistic activities of 2-alkyl-2-aminoadamantanes and analogues. Bioorg Med Chem Lett. 2008 Dec 1;18(23):6156-60. [CrossRef]
- Vasilevska, V., Guest, P.C., Bernstein, HG. et al. Molecular mimicry of NMDA receptors may contribute to neuropsychiatric symptoms in severe COVID-19 cases. J Neuroinflammation 18, 245 (2021). [CrossRef]
- Clarke G, Stilling RM, Kennedy PJ, Stanton C, Cryan JF, Dinan TG. Minireview: Gut microbiota: The neglected endocrine organ. Mol Endocrinol. 2014 Aug;28(8):1221-38. [CrossRef]
- Brown JM, Hazen SL. The gut microbial endocrine organ: Bacterially derived signals driving cardiometabolic diseases. Annu Rev Med. 2015;66:343-59. [CrossRef]
- Mu Q, Tavella VJ, Luo XM. Role of Lactobacillus reuteri in Human Health and Diseases. Front Microbiol. 2018 Apr 19;9:757. [CrossRef]
- Du T, Lei A, Zhang N, Zhu C. The Beneficial Role of Probiotic Lactobacillus in Respiratory Diseases. Front Immunol. 2022 May 31;13:908010. [CrossRef]
- Danhof HA, Lee J, Thapa A, Britton RA, Di Rienzi SC. Microbial stimulation of oxytocin release from the intestinal epithelium via secretin signaling. Gut Microbes. 2023 Dec;15(2):2256043. [CrossRef]
- Sheitman BB, Knable MB, Jarskog LF, Chakos M, Boyce LH, Early J, Lieberman JA. Secretin for refractory schizophrenia. Schizophr Res. 2004 Feb 1;66(2-3):177-81. PMID: 15061251. [CrossRef]
- Acevedo-Rodriguez A, Mani SK, Handa RJ. Oxytocin and Estrogen Receptor β in the Brain: An Overview. Front Endocrinol (Lausanne). 2015 Oct 15;6:160. [CrossRef]
- Wiegand V, Gimpl G. Specification of the cholesterol interaction with the oxytocin receptor using a chimeric receptor approach. Eur J Pharmacol. 2012 Feb 15;676(1-3):12-9. [CrossRef]
- Omolaoye, T.S.; Halabi, M.O.; Mubarak, M.; Cyril, A.C.; Duvuru, R.; Radhakrishnan, R.; Du Plessis, S.S. Statins and Male Fertility: Is There a Cause for Concern? Toxics 2022, 10, 627. [CrossRef]
- Di Stasi SL, MacLeod TD, Winters JD, Binder-Macleod SA. Effects of statins on skeletal muscle: A perspective for physical therapists. Phys Ther. 2010 Oct;90(10):1530-42. [CrossRef]
- Morofuji Y, Nakagawa S, Ujifuku K, Fujimoto T, Otsuka K, Niwa M, Tsutsumi K. Beyond Lipid-Lowering: Effects of Statins on Cardiovascular and Cerebrovascular Diseases and Cancer. Pharmaceuticals (Basel). 2022 Jan 26;15(2):151. [CrossRef]
- Yuta Aoki, Hidenori Yamasue, Reply: Does imitation act as an oxytocin nebulizer in autism spectrum disorder?, Brain, Volume 138, Issue 7, July 2015, Page e361. [CrossRef]
- Sridharan D, Levitin DJ, Menon V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc Natl Acad Sci U S A. 2008 Aug 26;105(34):12569-74. [CrossRef]
- Svalastog AL, Martinelli L. Representing life as opposed to being: The bio-objectification process of the HeLa cells and its relation to personalized medicine. Croat Med J. 2013 Aug;54(4):397-402. [CrossRef]
- Han, Y., Kim, S.Y. Endothelial senescence in vascular diseases: Current understanding and future opportunities in senotherapeutics. Exp Mol Med 55, 1–12 (2023). [CrossRef]
- Morioka S, Maueröder C, Ravichandran KS. Living on the Edge: Efferocytosis at the Interface of Homeostasis and Pathology. Immunity. 2019 May 21;50(5):1149-1162. [CrossRef]
- Papanastasiou E, Gaughran F, Smith S. Schizophrenia as segmental progeria. J R Soc Med. 2011 Nov;104(11):475-84. [CrossRef]
- Stevenson JR, McMahon EK, Boner W, Haussmann MF. Oxytocin administration prevents cellular aging caused by social isolation. Psychoneuroendocrinology. 2019 May;103:52-60. [CrossRef]
- Wang SC, Zhang F, Zhu H, Yang H, Liu Y, Wang P, Parpura V, Wang YF. Potential of Endogenous Oxytocin in Endocrine Treatment and Prevention of COVID-19. Front Endocrinol (Lausanne). 2022 May 3;13:799521. [CrossRef]
- Welch MG, Margolis KG, Li Z, Gershon MD. Oxytocin regulates mice's gastrointestinal motility, inflammation, macromolecular permeability, and mucosal maintenance. Am J Physiol Gastrointest Liver Physiol. 2014;307(open in a new window)(8(open in a new window)):G848–62. [CrossRef]
- Yan L, Sun X, Wang Z, Song M, Zhang Z. Regulation of social behaviors by p-Stat3 via oxytocin and its receptor in the nucleus accumbens of male Brandt's voles (Lasiopodomys brandtii). Horm Behav. 2020 Mar;119:104638. [CrossRef]
- An, X., Sun, X., Hou, Y. et al. Protective effect of oxytocin on LPS-induced acute lung injury in mice. Sci Rep 9, 2836 (2019). [CrossRef]
- Zhang, Y., Catts, V., Sheedy, D. et al. Cortical grey matter volume reduction in people with schizophrenia is associated with neuro-inflammation. Transl Psychiatry 6, e982 (2016). [CrossRef]
- Lewis DA, Lieberman JA. Catching up on schizophrenia: Natural history and neurobiology. Neuron. 2000 Nov;28(2):325-34. PMID: 11144342. [CrossRef]
- Hazlett EA, Buchsbaum MS, Haznedar MM, Newmark R, Goldstein KE, Zelmanova Y, Glanton CF, Torosjan Y, New AS, Lo JN, Mitropoulou V, Siever LJ. Cortical gray and white matter volume in unmedicated schizotypal and schizophrenia patients. Schizophr Res. 2008 Apr;101(1-3):111-23. [CrossRef]
- Ho B, Andreasen NC, Ziebell S, Pierson R, Magnotta V. Long-term Antipsychotic Treatment and Brain Volumes: A Longitudinal Study of First-Episode Schizophrenia. Arch Gen Psychiatry. 2011;68(2):128–137. [CrossRef]
- Momenabadi, S., Vafaei, A.A., Bandegi, A.R. et al. Oxytocin Reduces Brain Injury and Maintains Blood-Brain Barrier Integrity After Ischemic Stroke in Mice. Neuromol Med 22, 557–571 (2020). [CrossRef]
- Saffari M, Momenabadi S, Vafaei AA, Vakili A, Zahedi-Khorasani M. Prophylactic effect of intranasal oxytocin on brain damage and neurological disorders in global cerebral ischemia in mice. Iran J Basic Med Sci. 2021 Jan;24(1):79-84. PMID: 33643574. [CrossRef]
- Holm, M.; Taipale, H.; Tanskanen, A.; Tiihonen, J.; Mitterdorfer-Rutz, E. Employment among people with schizophrenia or bipolar disorder: A population-based study using nationwide registers. Acta Psychiatr. Scand. 2020, 143, 61–71. [Google Scholar] [CrossRef] [PubMed].
- Lévesque, I.S.; Abdel-Baki, A. Homeless youth with first-episode psychosis: A 2-year outcome study. Schizophr. Res. 2019, 216, 460–469. [Google Scholar] [CrossRef].
- Davidson, L.; Schmutte, T.; Dinzeo, T.; Andres-Hyman, R. Remission and Recovery in Schizophrenia: Practitioner and Patient Perspectives. Schizophr. Bull. 2007, 34, 5-8.
- Stone WS, Phillips MR, Yang LH, Kegeles LS, Susser ES, Lieberman JA. Neurodegenerative model of schizophrenia: Growing evidence to support a revisit. Schizophr Res. 2022 May;243:154-162. [CrossRef]
- Leucht, S.; Lasser, R. The Concepts of Remission and Recovery in Schizophrenia. Pharmacopsychiatry 2006, 39, 161–170.
- Liberman, R.P.; Kopelowicz, A.; Ventura, J.; Gutkind, D. Operational criteria and factors related to recovery from schizophrenia. Int. Rev. Psychiatry 2002, 14, 256–272.
- Leppien E, Mulcahy K, Demler TL, Trigoboff E, Opler L. Effects of Statins and Cholesterol on Patient Aggression: Is There a Connection? Innov Clin Neurosci. 2018 Apr 1;15(3-4):24-27.
- B.A. Golomb, T. Kane, J.E. Dimsdale, Severe irritability associated with statin cholesterol-lowering drugs, QJM: An International Journal of Medicine, Volume 97, Issue 4, April 2004, Pages 229–235. [CrossRef]
- Jacobo-Albavera L, Domínguez-Pérez M, Medina-Leyte DJ, González-Garrido A, Villarreal-Molina T. The Role of the ATP-Binding Cassette A1 (ABCA1) in Human Disease. Int J Mol Sci. 2021 Feb 5;22(4):1593. [CrossRef]
- Every-Palmer S, Souter V. Low vitamin D levels found in 95 % of psychiatric inpatients within a forensic service. Australian & New Zealand Journal of Psychiatry. 2015;49(4):391-392. [CrossRef]
- De Berardis D, Marini S, Piersanti M, Cavuto M, Perna G, Valchera A, Mazza M, Fornaro M, Iasevoli F, Martinotti G, Di Giannantonio M. The Relationships between Cholesterol and Suicide: ASen P, Adewusi D, Blakemore AI, Kumari V. How do lipids influence risk of violence, self-harm and suicidality in people with psychosis? A systematic review. Aust N Z J Psychiatry. 2022 May;56(5):451-488. n Update. ISRN Psychiatry. 2012 Dec 23;2012:387901. https://doi.org/10.5402/2012/387901. [CrossRef]
- Buydens-Branchey L, Branchey M, Hudson J, Fergeson P. Low HDL cholesterol, aggression and altered central serotonergic activity. Psychiatry Res. 2000 Mar 6;93(2):93-102. [CrossRef]
- Sahebzamani, Frances M. et al.Relationship among low cholesterol levels, depressive symptoms, aggression, hostility, and cynicism. Journal of Clinical Lipidology, Volume 7, Issue 3, 208 – 216.
- Hofhansel L, Weidler C, Votinov M, Clemens B, Raine A, Habel U. Morphology of the criminal brain: Gray matter reductions are linked to antisocial behavior in offenders. Brain Struct Funct. 2020 Sep;225(7):2017-2028. [CrossRef]
- Cristofori I, Zhong W, Mandoske V, Chau A, Krueger F, Strenziok M, Grafman J. Brain Regions Influencing Implicit Violent Attitudes: A Lesion-Mapping Study. J Neurosci. 2016 Mar 2;36(9):2757-68. [CrossRef]
- Pardini DA, Raine A, Erickson K, Loeber R. Lower amygdala volume in men is associated with childhood aggression, early psychopathic traits, and future violence. Biol Psychiatry. 2014 Jan 1;75(1):73-80. [CrossRef]
- Zhang R, Jiang X, Chang M, Wei S, Tang Y, Wang F. White matter abnormalities of corpus callosum in patients with bipolar disorder and suicidal ideation. Ann Gen Psychiatry. 2019 Sep 10;18:20. [CrossRef]
- Naudts K, Hodgins S. Neurobiological correlates of violent behavior among persons with schizophrenia. Schizophr Bull. 2006 Jul;32(3):562-72. [CrossRef]
- Yan R, Wei D, Varshneya A, Shan L, Dai B, Asencio HJ 3rd, Gollamudi A, Lin D. The multi-stage plasticity in the aggression circuit underlying the winner effect. Cell. 2024 Oct 9:S0092-8674(24)01088-2. [CrossRef]
- Popescu ER, Semeniuc S, Hritcu LD, Horhogea CE, Spataru MC, Trus C, Dobrin RP, Chirita V, Chirita R. Cortisol and Oxytocin Could Predict Covert Aggression in Some Psychotic Patients. Medicina (Kaunas). 2021 Jul 27;57(8):760. [CrossRef]
- Pfundmair M, Reinelt A, DeWall CN, Feldmann L. Oxytocin strengthens the link between provocation and aggression among low anxiety people. Psychoneuroendocrinology. 2018 Jul;93:124-132. [CrossRef]
- DeWall, C. N., Gillath, O., Pressman, S. D., Black, L. L., Bartz, J. A., Moskovitz, J., & Stetler, D. A. (2014). When the Love Hormone Leads to Violence: Oxytocin Increases Intimate Partner Violence Inclinations Among High Trait Aggressive People. Social Psychological and Personality Science, 5(6), 691-697. [CrossRef]
- Heitmann A.S.B., Zanjani A.A.H., Klenow M.B., Mularski A., Sønder S.L., Lund F.W., Boye T.L., Dias C., Bendix P.M., Simonsen A.C.; et al. Phenothiazines alter plasma membrane properties and sensitize cancer cells to injury by inhibiting annexin-mediated repair. J. Biol. Chem. 2021;297:101012. [CrossRef]
- Voronova, O.; Zhuravkov, S.; Korotkova, E.; Artamonov, A.; Plotnikov, E. Antioxidant Properties of New Phenothiazine Derivatives. Antioxidants 2022, 11, 1371. [CrossRef]
- Keynes RG, Karchevskaya A, Riddall D, Griffiths CH, Bellamy TC, Chan AWE, Selwood DL, Garthwaite J. N10 -carbonyl-substituted phenothiazines inhibiting lipid peroxidation and associated nitric oxide consumption powerfully protect brain tissue against oxidative stress. Chem Biol Drug Des. 2019 Sep;94(3):1680-1693. [CrossRef]
- Mehrabi SF, Elmi S, Nylandsted J. Repurposing phenothiazines for cancer therapy: Compromising membrane integrity in cancer cells. Front Oncol. 2023 Nov 23;13:1320621. [CrossRef]
- Amaral L, Viveiros M, Molnar J. Antimicrobial activity of phenothiazines. In Vivo. 2004 Nov-Dec;18(6):725-31.
- Zhang X., Meng F., Lyu W., He J., Wei R., Du Z., Zhang C. Histone lactylation antagonizes senescence and skeletal muscle aging via facilitating gene expression reprogramming. bioRxiv. 2023. [CrossRef]
- Wei L., Yang X., Wang J., Wang Z., Wang Q., Ding Y., Yu A. H3K18 lactylation of senescent microglia potentiates brain aging and Alzheimer’s disease through the NFκB signaling pathway. J. Neuroinflamm. 2023;20:208. [CrossRef]
- Munteanu C, Schwartz B. The relationship between nutrition and the immune system. Front Nutr. 2022 Dec 8;9:1082500. [CrossRef]
- Link C.D. Is There a Brain Microbiome? Neurosci. Insights. 2021;16:26331055211018709. [CrossRef]
- Pretorius L., Kell D.B., Pretorius E. Iron Dysregulation and Dormant Microbes as Causative Agents for Impaired Blood Rheology and Pathological Clotting in Alzheimer’s Type Dementia. Front. Neurosci. 2018;12:851. [CrossRef]
- Mahin Ghorbani, Unveiling the Human Brain Virome in Brodmann Area 46: Novel Insights Into Dysbiosis and Its Association With Schizophrenia, Schizophrenia Bulletin Open, Volume 4, Issue 1, January 2023, sgad029. [CrossRef]
- Safe S, Jin UH, Park H, Chapkin RS, Jayaraman A. Aryl Hydrocarbon Receptor (AHR) Ligands as Selective AHR Modulators (SAhRMs). Int J Mol Sci. 2020 Sep 11;21(18):6654. [CrossRef]
- Sharma R. Emerging Interrelationship between the Gut Microbiome and Cellular Senescence in the Context of Aging and Disease: Perspectives and Therapeutic Opportunities. Probiotics Antimicrob. Proteins. 2022;14:648–663. [CrossRef]
- Velázquez-Marrero, Cristina, and José R. Lemos. 2024. ‘Modulation of Oxytocin Release by Internal Calcium Stores.’ Oxytocin and Social Function. IntechOpen. [CrossRef]
- Torti MF, Giovannoni F, Quintana FJ, García CC. The Aryl Hydrocarbon Receptor as a Modulator of Anti-viral Immunity. Front Immunol. 2021 Mar 5;12:624293. [CrossRef]
- Kueck T, Cassella E, Holler J, Kim B, Bieniasz PD. The aryl hydrocarbon receptor and interferon gamma generate antiviral states via transcriptional repression. Elife. 2018 Aug 22;7:e38867. [CrossRef]
- Ren J, Lu Y, Qian Y, Chen B, Wu T, Ji G. Recent progress regarding kaempferol for treating various diseases. Exp Ther Med. 2019 Oct;18(4):2759-2776. [CrossRef]
- Xia QS, Wu F, Wu WB, Dong H, Huang ZY, Xu L, Lu FE, Gong J. Berberine reduces hepatic ceramide levels to improve insulin resistance in HFD-fed mice by inhibiting HIF-2α. Biomed Pharmacother. 2022 Jun;150:112955. [CrossRef]
- Imran M, Ghorat F, Ul-Haq I, Ur-Rehman H, Aslam F, Heydari M, Shariati MA, Okuskhanova E, Yessimbekov Z, Thiruvengadam M, Hashempur MH, Rebezov M. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants (Basel). 2020 Aug 4;9(8):706. [CrossRef]
- Kim TW. Fisetin, an Anti-Inflammatory Agent, Overcomes Radioresistance by Activating the PERK-ATF4-CHOP Axis in Liver Cancer. Int J Mol Sci. 2023 May 22;24(10):9076. [CrossRef]
- Lamming DW. Inhibition of the Mechanistic Target of Rapamycin (mTOR)-Rapamycin and Beyond. Cold Spring Harb Perspect Med. 2016 May 2;6(5):a025924. [CrossRef]
- Zamani M, Nikbaf-Shandiz M, Aali Y, Rasaei N, Zarei M, Shiraseb F, Asbaghi O. The effects of acarbose treatment on cardiovascular risk factors in impaired glucose tolerance and diabetic patients: A systematic review and dose-response meta-analysis of randomized clinical trials. Front Nutr. 2023 Aug 1;10:1084084. [CrossRef]
- Singh CK, Chhabra G, Ndiaye MA, Garcia-Peterson LM, Mack NJ, Ahmad N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid Redox Signal. 2018 Mar 10;28(8):643-661. [CrossRef]
- Janić M, Lunder M, Cerkovnik P, Prosenc Zmrzljak U, Novaković S, Šabovič M. Low-Dose Fluvastatin and Valsartan Rejuvenate the Arterial Wall Through Telomerase Activity Increase in Middle-Aged Men. Rejuvenation Res. 2016 Apr;19(2):115-9. [CrossRef]
- Kuk MU, Kim JW, Lee YS, Cho KA, Park JT, Park SC. Alleviation of Senescence via ATM Inhibition in Accelerated Aging Models. Mol Cells. 2019 Mar 31;42(3):210-217. [CrossRef]
- S. An, S. Cho, J. Kang, S. Lee, H. Kim, D. Min, E. Son, K. Cho, Inhibition of 3-phosphoinositide–dependent protein kinase 1 (PDK1) can revert cellular senescence in human dermal fibroblasts, Proc. Natl. Acad. Sci. U.S.A. 117 (49) 31535-31546. (2020). [CrossRef]
- Sheitman BB, Knable MB, Jarskog LF, Chakos M, Boyce LH, Early J, Lieberman JA. Secretin for refractory schizophrenia. Schizophr Res. 2004 Feb 1;66(2-3):177-81. PMID: 15061251. [CrossRef]
- Danhof HA, Lee J, Thapa A, Britton RA, Di Rienzi SC. Microbial stimulation of oxytocin release from the intestinal epithelium via secretin signaling. Gut Microbes. 2023 Dec;15(2):2256043. [CrossRef]
- Wu, Y., Zhang, X., Liu, X. et al. Galactooligosaccharides and Limosilactobacillus reuteri synergistically alleviate gut inflammation and barrier dysfunction by enriching Bacteroides acidifaciens for pentadecanoic acid biosynthesis. Nat Commun 15, 9291 (2024). [CrossRef]
- Okubo R, Koga M, Katsumata N, Odamaki T, Matsuyama S, Oka M, Narita H, Hashimoto N, Kusumi I, Xiao J, Matsuoka YJ. Effect of bifidobacterium breve A-1 on anxiety and depressive symptoms in schizophrenia: A proof-of-concept study. J Affect Disord. 2019 Feb 15;245:377-385. [CrossRef]
- Sfera, A.; Thomas, K.A.; Anton, J. Cytokines and Madness: A Unifying Hypothesis of Schizophrenia Involving Interleukin-22. Int. J. Mol. Sci. 2024, 25, 12110. [CrossRef]
- Van Spaendonk H, Ceuleers H, Witters L, Patteet E, Joossens J, Augustyns K, Lambeir AM, De Meester I, De Man JG, De Winter BY. Regulation of intestinal permeability: The role of proteases. World J Gastroenterol. 2017 Mar 28;23(12):2106-2123. [CrossRef]
- Paramos-de-Carvalho D, Jacinto A, Saúde L. The right time for senescence. Elife. 2021 Nov 10;10:e72449. [CrossRef]
- 85. Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: Senolytics and senomorphics. FEBS J. 2023 Mar;290(5):1362-1383. [CrossRef]
- Yue L, Ren Y, Yue Q, Ding Z, Wang K, Zheng T, Chen G, Chen X, Li M, Fan L. α-Lipoic Acid Targeting PDK1/NRF2 Axis Contributes to the Apoptosis Effect of Lung Cancer Cells. Oxid Med Cell Longev. 2021 Jun 4;2021:6633419. [CrossRef]
- Rossell SL, Francis PS, Galletly C, Harris A, Siskind D, Berk M, Bozaoglu K, Dark F, Dean O, Liu D, Meyer D, Neill E, Phillipou A, Sarris J, Castle DJ. N-acetylcysteine (NAC) in schizophrenia resistant to clozapine: A double blind randomised placebo controlled trial targeting negative symptoms. BMC Psychiatry. 2016 Sep 15;16(1):320. [CrossRef]
- 89. Sanders LLO, de Souza Menezes CE, Chaves Filho AJM, de Almeida Viana G, Fechine FV, Rodrigues de Queiroz MG, Gonçalvez da Cruz Fonseca S, Mendes Vasconcelos SM, Amaral de Moraes ME, Gama CS, Seybolt S, de Moura Campos E, Macêdo D, Freitas de Lucena D. α-Lipoic Acid as Adjunctive Treatment for Schizophrenia: An Open-Label Trial. J Clin Psychopharmacol. 2017 Dec;37(6):697-701. [CrossRef]
- Spellmann I, Rujescu D, Musil R, Meyerwas S, Giegling I, Genius J, Zill P, Dehning S, Cerovecki A, Seemüller F, Schennach R, et al/. Pleckstrin homology domain containing 6 protein (PLEKHA6) polymorphisms are associated with psychopathology and response to treatment in schizophrenic patients. Prog Neuropsychopharmacol Biol Psychiatry. 2014 Jun 3;51:190-5. [CrossRef]
- Feng L, Wong JCM, Mahendran R, Chan ESY, Spencer MD. Intranasal oxytocin for autism spectrum disorders (ASD). Cochrane Database Syst Rev. 2017 Jan 20;2017(1): CD010928. [CrossRef]
- Nicolson GL, Ash ME. Membrane Lipid Replacement for chronic illnesses, aging and cancer using oral glycerolphospholipid formulations with fructooligosaccharides to restore phospholipid function in cellular membranes, organelles, cells and tissues. Biochim Biophys Acta Biomembr. 2017 Sep;1859(9 Pt B):1704-1724. [CrossRef]
- Repetto, Marisa, Jimena Semprine, and Alberto Boveris. 2012. ‘Lipid Peroxidation: Chemical Mechanism, Biological Implications and Analytical Determination’. Lipid Peroxidation. InTech. [CrossRef]
- Tatsuya Sato, Jason Solomon, ShapiroHsiang-Chun, Chang Richard A, Miller Hossein Ardehali (2022) Aging is associated with increased brain iron through cortex-derived hepcidin expression eLife 11:e73456. [CrossRef]
- Lolicato F, Juhola H, Zak A, Postila PA, Saukko A, Rissanen S, Enkavi G, Vattulainen I, Kepczynski M, Róg T. Membrane-Dependent Binding and Entry Mechanism of Dopamine into Its Receptor. ACS Chem Neurosci. 2020 Jul 1;11(13):1914-1924. [CrossRef]
- Dichtl S, Demetz E, Haschka D, Tymoszuk P, Petzer V, Nairz M, Seifert M, Hoffmann A, Brigo N, Würzner R, Theurl I, Karlinsey JE, Fang FC, Weiss G. Dopamine Is a Siderophore-Like Iron Chelator That Promotes Salmonella enterica Serovar Typhimurium Virulence in Mice. mBio. 2019 Feb 5;10(1):e02624-18. [CrossRef]
- Tao L, Qing Y, Cui Y, Shi D, Liu W, Chen L, Cao Y, Dai Z, Ge X, Zhang L. Lysosomal membrane permeabilization mediated apoptosis involve in perphenazine-induced hepatotoxicity in vitro and in vivo. Toxicol Lett. 2022 Aug 15;367:76-87. [CrossRef]
- Yacoub A, Park MA, Hanna D, Hong Y, Mitchell C, Pandya AP, Harada H, Powis G, Chen CS, Koumenis C, Grant S, Dent P. OSU-03012 promotes caspase-independent but PERK-, cathepsin B-, BID-, and AIF-dependent killing of transformed cells. Mol Pharmacol. 2006 Aug;70(2):589-603. [CrossRef]
- Varalda M, Antona A, Bettio V, Roy K, Vachamaram A, Yellenki V, Massarotti A, Baldanzi G, Capello D. Psychotropic Drugs Show Anticancer Activity by Disrupting Mitochondrial and Lysosomal Function. Front Oncol. 2020 Oct 19;10:562196. [CrossRef]
- Girgis RR, Lieberman JA. Anti-viral properties of antipsychotic medications in the time of COVID-19. Psychiatry Res. 2021 Jan;295:113626. [CrossRef]
- Zhang S, Suvannasankha A, Crean CD, White VL, Johnson A, Chen CS, Farag SS. OSU-03012, a novel celecoxib derivative, is cytotoxic to myeloma cells and acts through multiple mechanisms. Clin Cancer Res. 2007 Aug 15;13(16):4750-8. [CrossRef]
- Tsao N, Chang YC, Hsieh SY, Li TC, Chiu CC, Yu HH, Hsu TC, Kuo CF. AR-12 Has a Bactericidal Activity and a Synergistic Effect with Gentamicin against Group A Streptococcus. Int J Mol Sci. 2021 Oct 27;22(21):11617. [CrossRef]
- Nehme H, Saulnier P, Ramadan AA, Cassisa V, Guillet C, Eveillard M, Umerska A. Antibacterial activity of antipsychotic agents, their association with lipid nanocapsules and its impact on the properties of the nanocarriers and on antibacterial activity. PLoS ONE. 2018 Jan 3;13(1):e0189950. [CrossRef]
- Girgis RR, Lieberman JA. Anti-viral properties of antipsychotic medications in the time of COVID-19. Psychiatry Res. 2021 Jan;295:113626. [CrossRef]
- Roberts RA, Kim J, Poklepovic A, Roberts JL, Booth L, Dent P. AR12 (OSU-03012) suppresses GRP78 expression and inhibits SARS-CoV-2 replication. Biochem Pharmacol. 2020 Dec;182:114227. [CrossRef]
- Zhang S, Suvannasankha A, Crean CD, White VL, Johnson A, Chen CS, Farag SS. OSU-03012, a novel celecoxib derivative, is cytotoxic to myeloma cells and acts through multiple mechanisms. Clin Cancer Res. 2007 Aug 15;13(16):4750-8. [CrossRef]
Figure 1.
IL-10 and IL-22 share their receptor (IL-10 receptor beta and IL-22 receptor 1). IL-22 attachment activates both STAT3 and MAPK. Decreased BDNF upregulates STAT3, linked to the cholinergic system and the brain, especially the insular cortex (IC) and anterior cingulate cortex (ACC).
Figure 1.
IL-10 and IL-22 share their receptor (IL-10 receptor beta and IL-22 receptor 1). IL-22 attachment activates both STAT3 and MAPK. Decreased BDNF upregulates STAT3, linked to the cholinergic system and the brain, especially the insular cortex (IC) and anterior cingulate cortex (ACC).
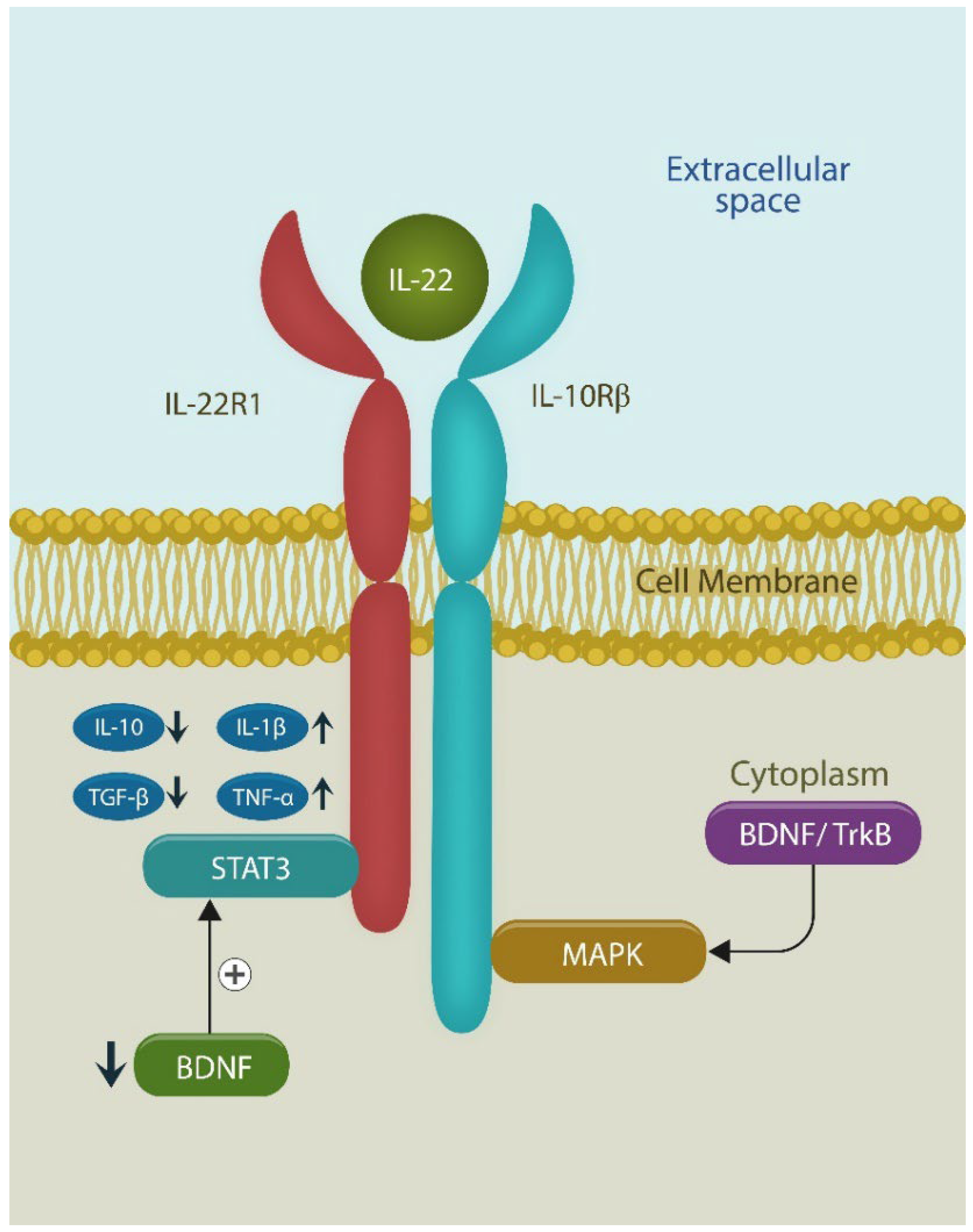
Figure 2.
SCZ starts in early childhood with a premorbid phase with no symptoms or mild ones. Attenuated symptoms, such as social isolation, anxiety, and insomnia, mark the prodromal phase. This phase blends gradually into psychosis, during which patients are usually hospitalized many times for exhibiting positive symptoms. Around midlife, the positive symptoms gradually subside and are replaced by negative and cognitive manifestations (figure adapted from Liberman).
Figure 2.
SCZ starts in early childhood with a premorbid phase with no symptoms or mild ones. Attenuated symptoms, such as social isolation, anxiety, and insomnia, mark the prodromal phase. This phase blends gradually into psychosis, during which patients are usually hospitalized many times for exhibiting positive symptoms. Around midlife, the positive symptoms gradually subside and are replaced by negative and cognitive manifestations (figure adapted from Liberman).
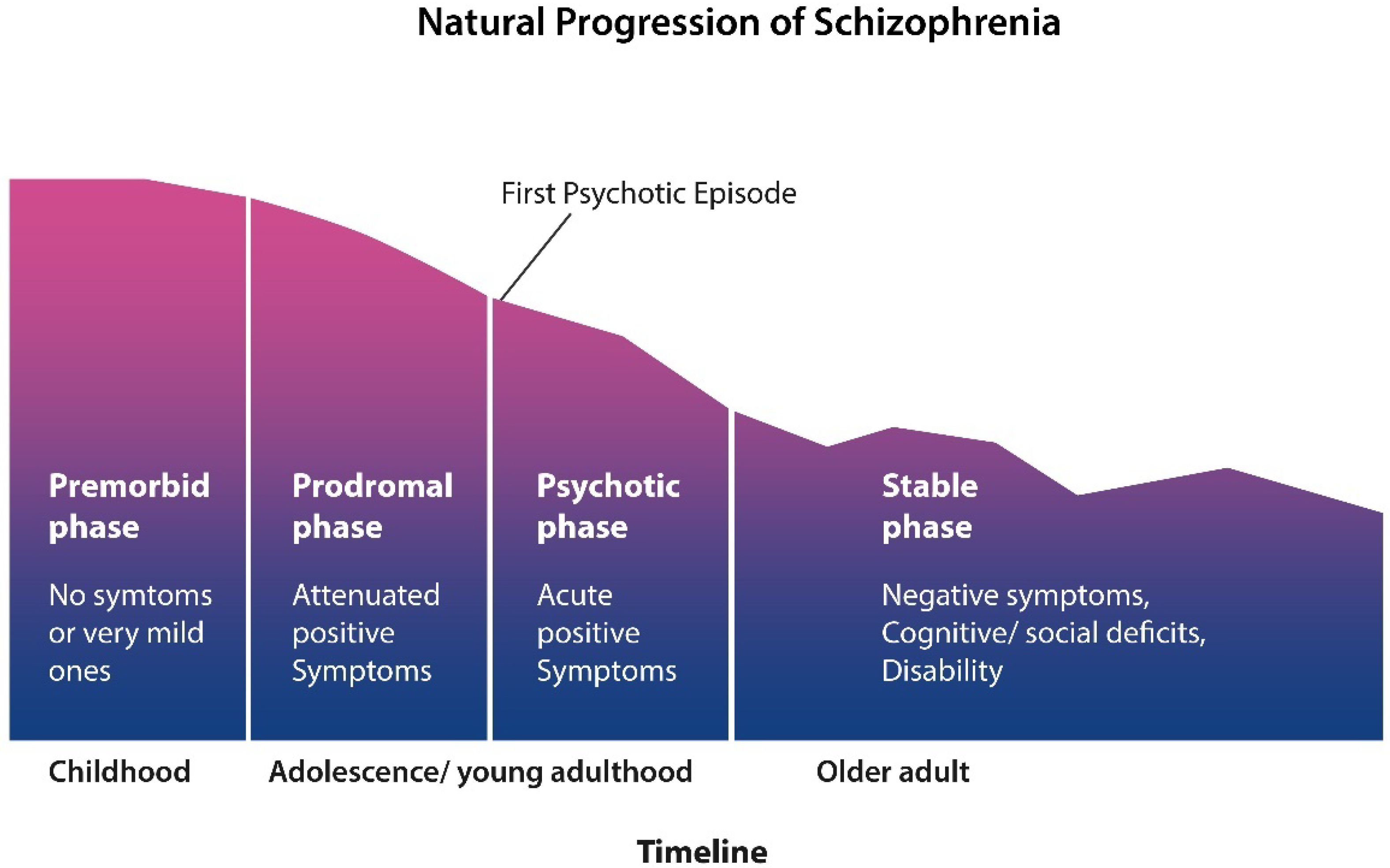
Figure 3.
Schematic representation of the SARS-CoV-2 virus antigen S (spike) with the known repeats (motifs) mimicking human proteins. The triple arginine motif promotes pore-forming in cell membrane via its side chain. HAP2, or GCS1, is a family of membrane fusion proteins found in the sperm cells that are directly related to male fertility. Calcium calmodulin kinase II hijacks OXYTOCIN and promotes phosphorylation of Tau protein (found in both the brain and placenta). SNARE proteins participate in vesicular and synaptic transmission. Human endogenous retrovirus (envelope) sequences mimic placental syncytin, a trophoblast fusogen. NSP-1 is a non-S antigen of SARS-CoV-2 protein that resembles tubulin found in the CNS and placenta.
Figure 3.
Schematic representation of the SARS-CoV-2 virus antigen S (spike) with the known repeats (motifs) mimicking human proteins. The triple arginine motif promotes pore-forming in cell membrane via its side chain. HAP2, or GCS1, is a family of membrane fusion proteins found in the sperm cells that are directly related to male fertility. Calcium calmodulin kinase II hijacks OXYTOCIN and promotes phosphorylation of Tau protein (found in both the brain and placenta). SNARE proteins participate in vesicular and synaptic transmission. Human endogenous retrovirus (envelope) sequences mimic placental syncytin, a trophoblast fusogen. NSP-1 is a non-S antigen of SARS-CoV-2 protein that resembles tubulin found in the CNS and placenta.
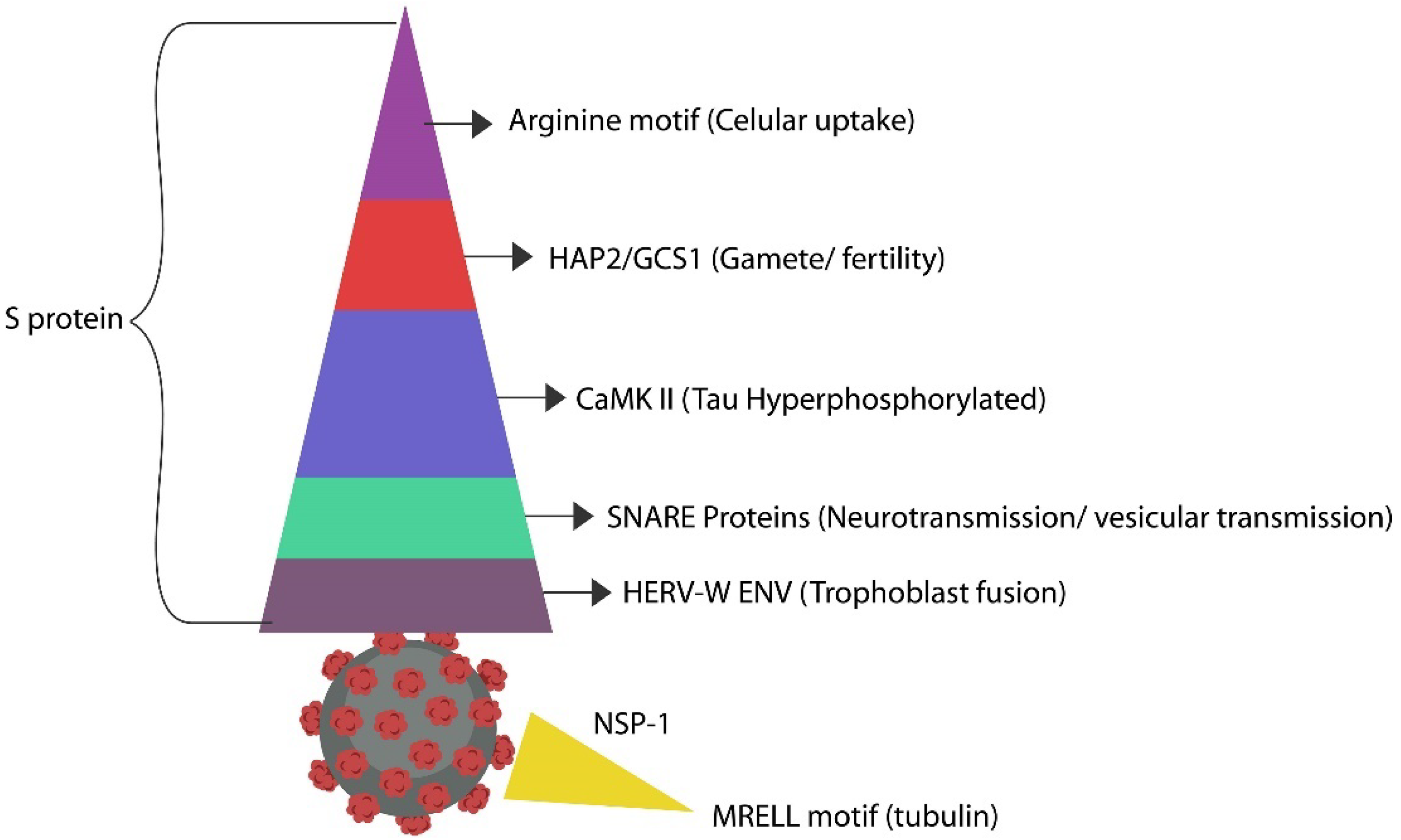
Figure 4.
Upon growth factor-receptor binding, PDK-1 is recruited to the cell membrane and activated by PI3K via PIP1 and PIP2. MLR, especially the glycerophospholipid phosphatidylserine (PS), inhibits PDK1 by maintaining inactive conformation. Downstream, Akt phosphorylation is inhibited, releasing the inhibitory break from GSK-3β (lack of inhibitory phosphorylation). Together, phosphorylation changes contribute to neuropathology, including SCZ. Some second-generation antipsychotic drugs upregulate GSK-3β (by increasing activating phosphorylation). OSU-03012 acts synergistically with MLR, inhibiting PDK1.
Figure 4.
Upon growth factor-receptor binding, PDK-1 is recruited to the cell membrane and activated by PI3K via PIP1 and PIP2. MLR, especially the glycerophospholipid phosphatidylserine (PS), inhibits PDK1 by maintaining inactive conformation. Downstream, Akt phosphorylation is inhibited, releasing the inhibitory break from GSK-3β (lack of inhibitory phosphorylation). Together, phosphorylation changes contribute to neuropathology, including SCZ. Some second-generation antipsychotic drugs upregulate GSK-3β (by increasing activating phosphorylation). OSU-03012 acts synergistically with MLR, inhibiting PDK1.
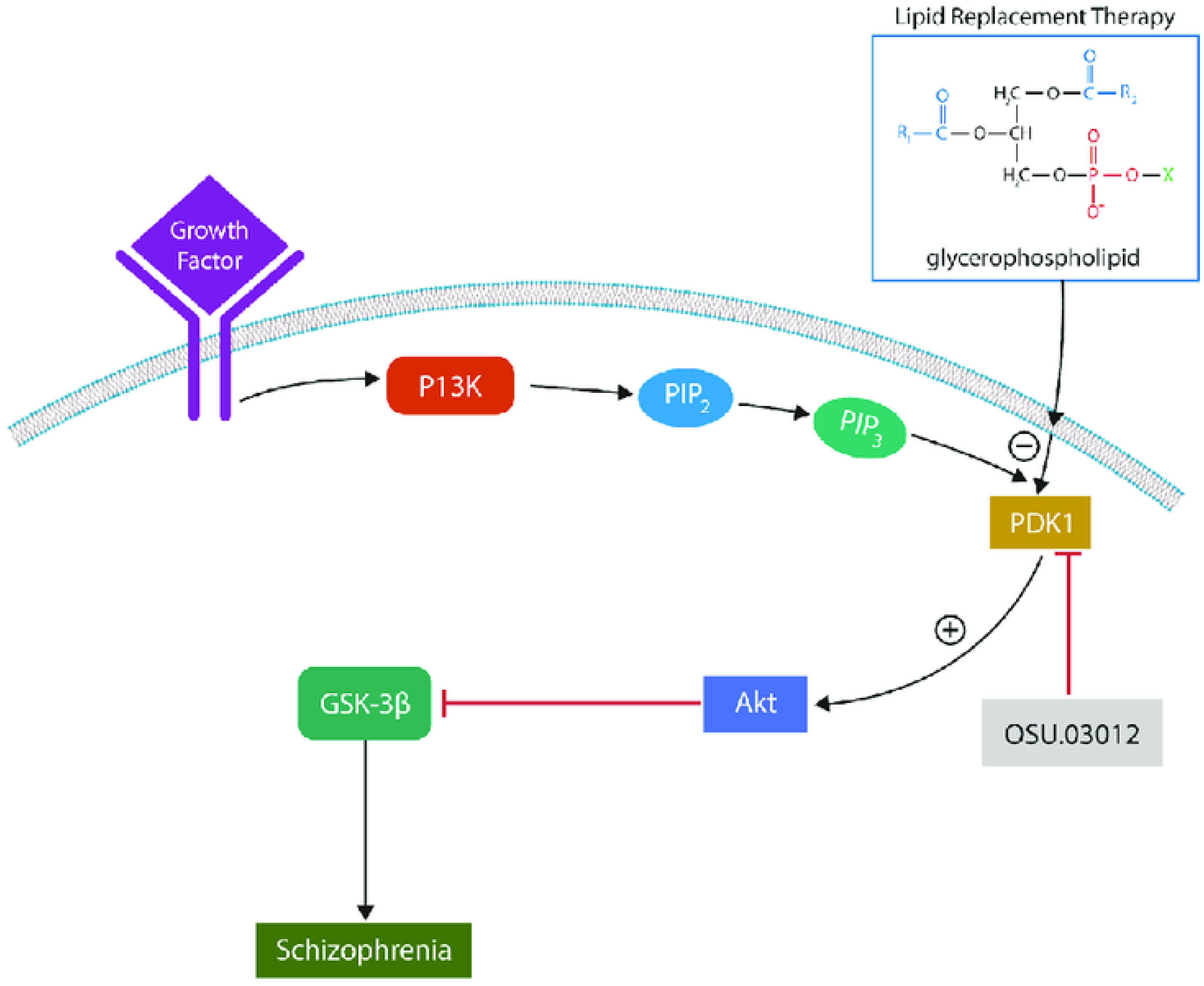
Table 2.
OSU 03012 and antipsychotic drugs, common characteristic.
| OSU-03012 | Antipsychotic drugs | References |
|---|---|---|
| Lowers ER stress via PERK | Lower ER stress via PERK | 106 |
| Inhibits cathepsins | Inhibits cathepsins | 107; 108 |
| Antineoplastic properties | Antineoplasic properties | 109;110 |
| Antibacterial properties | Antibacterial properties | 111, 112 |
| Antiviral properties | Antiviral properties | 113;114 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



