
Submitted:
04 July 2024
Posted:
04 July 2024
You are already at the latest version
Abstract
Epilepsy is a non-communicable neurological disease characterized by abnormal brain activity with conceivable peripheral implications. Few studies examine the role of peripheral and brain inflammation in the relationship between seizure predisposition and nonalcoholic fatty liver disease. Severe early-life infections leading to sepsis results in increased inflammation that can aggravate epilepsy and hepatic damage progression both associated with increased odds for hospital admissions in epileptic patients. Here, we examined the effect of early LPS challenge (1 mg/kg, i.p. at post-natal day3, PND3) in inducing hepatic damage in a genetic model of young epileptic WAG/Rij rats (PND45), evaluating the intra- and inter-gender differences on systemic and liver inflammation, hepatic lipid dysmetabolism and oxidative damage associated to mitochondrial functional impairment. First, epileptic rats exposed to LPS, regardless of gender, displayed increased serum hepatic enzymes and altered lipid profile. Early LPS challenge induced a major inflammatory and immune response in male epileptic rats than females in both serum and liver, increasing pro-inflammatory cytokines and hepatic immune cell recruitment. Conversely, LPS-insulted females showed a marked alteration in hepatic metabolic and lipid profile and the reduction in mitochondrial fatty acid oxidation. The two different gender-related mechanisms of LPS-induced liver injury converge in mitochondrial oxidative damage with intensified ROS production in both sexes, that notably induced a compensatory increase in antioxidant defense only in females. Our study with a translational potential points out that early postnatal infections predispose epileptic rats to develop or worsen hepatic disorders in a sex-dependent manner, amplifying inflammation, lipid dysmetabolism, and mitochondrial impairment.
Keywords:
genetic animal model
; epilepsy
; mitochondrial bioenergetics
; oxidative damage
; sex-dependent alterations
; neonatal infections
1. Introduction
The liver is the crucial organ involved in both immunity and metabolism in children and adults [1]. However, how detrimental bacterial challenges in the post-natal period can influence liver adaptation and immune function is overlooked. The hepatic immune response is involved in the systemic response to severe infection and is mainly orchestrate by Kupffer cells (KCs) [2,3]. These liver-resident macrophagic cells play a critical role in maintaining liver functions and are responsible for innate inflammatory and immune response, , along with recruited monocytes and neutrophils, and stimulating T cells presenting antigens [4]. KCs engulf and eliminate pathogens, as well as cell debris by phagocytosis and apoptotic cells, and function as active sensors that detect immunoreactive fragments, including lipopolysaccharide (LPS). The latter is an endotoxin constituting a structural part of the outer membrane of Gram-negative bacteria, capable of significantly stimulating the immune system [5] and therefore widely applied in the experimental models to resemble bacterial infections and sepsis [6]. Activated KCs secrete several mediators that regulate inflammation and homeostasis, as well as drive the inflammatory response to liver injury [7]. Consistently, hepatocytes and immune cells cooperate in controlling systemic and local bacterial infections [7].
However, mechanisms by which sepsis induce organ dysfunction and dysmetabolism have not been fully clarified. Mitochondrial alterations and reactive oxygen species (ROS) have been proposed in the pathogenesis of both neonatal and adult sepsis [8,9,10]. During sepsis, the liver plays two opposing roles: a source of inflammatory mediators and a target organ for a “boomerang effect” of the inflammatory mediators [6]. Consistently, neonatal rat endotoxemia is strictly linked to hepatic mitochondrial alterations [9].
Liver diseases, in both acute and chronic forms, can be associated with a wide spectrum of neurological manifestations, ranging in severity from subclinical changes to neurocritical conditions [11]. Among all, epilepsy is a non-communicable neurological disease characterized by abnormal brain activity with feasible peripheral implications. . Indeed, in epileptic patients with hepatic disease, meticulous concerns should be made regarding the choice of the appropriate anti-epileptic therapy.
In vivo studies have demonstrated that systemic and central nervous system inflammation is the main actor in the relationship between seizure predisposition and the development of nonalcoholic fatty liver disease (NAFLD) [12]. Indeed, severe early-life infections leading to sepsis may result in triggering hepatic and neuro-inflammation that can aggravate epilepsy. Furthermore, NAFLD has been recently identified as an independent risk factor for sepsis in a large clinical cohort showing a strong relationship between mortality in NAFLD-associated sepsis and hepatic mitochondrial and energetic metabolism dysfunction [13].
Growing lines of evidence have heightened the importance of investigating sex-based differences in medicine and research about different pathologies including infectious diseases [14,15]. Indeed, many bacterial infections show gender distinctions in pathophysiology, incidence, as well as clinical presentation, disease course, and outcome [14].
Here, we examined the hepatic detrimental effect of an early LPS challenge in young epileptic rats (WAG/Rij), pointing out the possible gender-related differences on liver inflammation and lipid metabolism alterations, associated to mitochondrial oxidative damage.
WAG/Rij rats, also named Wistar Albino Glaxo/Rijswijk rats, are a well-established animal model of human absence epilepsy, that share brain activity recording and many behavioral features resembling absence epilepsy in humans, including the translatable efficacy of different antiepileptic drugs [16], and their side effects, such as hepatotoxicity [17]. However, the characterization of the metabolic profile of this useful strain and its gender differences have been still unexplored.
2. Materials and Methods
2.1. In Vivo Experimental Procedures and Ethics Statement
All the experiments were carried out in male and female Wistar Albino Glaxo/Rijswijk (WAG/Rij) rat pups obtained after mating male epileptic rats with dams. At postnatal day (PND) 3, WAG/Rij pups received the single intraperitoneal injection of lipopolysaccharide (LPS, 1 mg/kg, Sigma-Aldrich) to mimic early immune activation by gram-negative infection. At PND45, the animals of both sexes were sacrificed, and serum samples and fresh or frozen liver tissue were collected for the following biochemical and molecular determinations. Since the limitations due to the impossibility to stereotaxically implanted 45-days-old rats for seizure quantification, preliminary results from electroencephalogram (EEG) recordings of 3-months-old male WAG/Rij rats challenged or not with LPS were obtained and reported in Supplementary Figure S1. All procedures involving the animals were carried out in conformity with international and national law and policies, including European Union (EU) Directive 2010/63/EU for animal experiments, Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines 2.0, https://www. arriveguidelines.org/resources, the Basel Declaration, and the National Centre for the Replacement, Refinement and Reduction of Animals in Research (NC3Rs) concept, and were approved by the Institutional Committee on the Ethics of Animal Experiments (CSV) of the University of Naples Federico II and by the Italian Ministry of Health under protocol No. 591/2020-PR.
2.2. Biochemical Evaluations of Serum Hepatic Parameters, Lipid Profile, and Inflammatory/Immune Mediators
At PND45, blood was collected from all experimental groups. Then, serum was obtained by centrifugation at 2500 rpm at 4 ◦C for 12 min and stored at -80◦C for subsequent biochemical analyses. Serum parameters (cholesterol, triglycerides, alanine aminotransferase or ALT, aspartate aminotransferase or AST, alkaline phosphatase or ALP, and lactate dehydrogenase or LDH) were measured using commercially available ELISA kits. Concomitantly, the concentration of twenty-three out of pro- and anti-inflammatory mediators, and factors of innate and adaptative immunity were obtained by a high sensitivity kit (Bio-Techne; R&D Systems, Inc., USA) using the Bio-Plex System and Luminex xMAP technology (Bio-Rad Laboratories, Inc., USA). Cytokine concentrations were derived by interpolating the measured fluorescence intensities to standard curves and correcting by the corresponding dilution factor employed to achieve the minimum volume for analysis. Bio-Plex Manager software was employed to calculate cytokine concentrations.
2.3. Mitochondrial Bioenergetics and Redox Status Evaluation
Hepatic mitochondrial isolation and oxidative capacities were performed as previously reported [17]. Oxygen consumption was polarographically measured using a Clark-type electrode in the presence of substrates and ADP (state 3) or with substrates alone (state 4). High quality of mitochondrial preparations was indicated by high respiratory control ratio values in all groups (data not shown), calculated as the ratio between states 3 and 4, according to Estabrook (1967). The degree of coupling was determined in the liver as previously reported [18] by applying equation by [19] degree of coupling = √(1-〖(Jo)〗_sh/〖(Jo)〗_unc ) where (Jo)sh represents the oxygen consumption rate in the presence of oligomycin that inhibits ATP synthase, and (Jo)unc is the uncoupled rate of oxygen consumption induced by carbonyl cyanide p-trifluoromethoxy phenylhydrazone (FCCP), which dissipates the trans-mitochondrial proton gradient. (Jo)sh and (Jo)unc were measured as above using succinate (10 mmol/L), rotenone (3.75 µmol/L) in the presence of oligomycin (2 µg/mL) or FCCP (1 µmol/L), respectively. The specific activity of the carnitine palmitoyltransferase (CPT) system and superoxide dismutase (SOD) was measured spectrophotometrically, as previously reported [20]. The rate of mitochondrial H2O2 release was assayed by measuring the linear increase in fluorescence caused by the oxidation of homovanillic acid in the presence of horseradish peroxidase. The protein content of the mitochondrial suspension was determined by the method of [21] using BSA as the protein standard. Furthermore, reduced glutathione (GSH) and oxidized glutathione (GSSG) concentrations in the liver were measured with the dithionitrobenzoic acid-GSSG reductase recycling assay [22]; the GSH-to-GSSG ratio was used as an oxidative stress marker.
2.4. ROS Assay
ROS assay was performed diluting an equal volume of freshly prepared tissue homogenate in 100 mM potassium phosphate buffer (pH 7.4) and a final concentration of 5 μM dichloro-fluorescein diacetate (Sigma-Aldrich, Milan, Italy) in dimethyl sulfoxide for 15 min at 37 ◦C was added. The dye loaded samples were centrifuged at 12,500× g per 10 min at 4 ◦C. The pellet was mixed at ice-cold temperatures in 5 mL of 100 mM potassium phosphate buffer (pH 7.4) and then incubated for 60 min at 37 ◦C. The fluorescence was measured by the HTS-7000 Plus-plate-reader spectrofluorometer (Perkin Elmer, Wellesley, MA, USA) at 488 nm for excitation and 525 nm for emission wavelengths. ROS were quantified from the dichloro-fluorescein standard curve in dimethyl sulfoxide (0–1 mM). For MDA assay, tissues were homogenized in 1.15% KCl solution.
2.5. RNA Extraction and Semi-Quantitative Real-Time (RT)-PCR
Total RNA isolated from the liver was extracted using TRIzol Reagent (Bio-Rad Laboratories, Hercules, CA, USA; 7326890) following the extraction kit’s protocol for RNA (NucleoSpin®, Macherey-Nagel GmbH & Co, Düren, Germany; FC140955N). cDNA was obtained using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA; 4374966) from 8 μg total RNA. RT-PCRs were performed with a Bio-Rad CFX96 Connect Real-time PCR System instrument and software (Bio-Rad Laboratories). The RT-PCR conditions were 15 min at 95°C followed by 40 cycles of two-step PCR denaturation at 94°C for 15 s, annealing extension at 55°C for 30 s and extension at 72°C for 30 s. Each sample contained 500 ng cDNA in 2X QuantiTect SYBR Green PCR Master Mix (204145) and primers pairs to amplify interleukin (IL)-1β (IL1b), cyclooxygenase-2 (PTGS2), tumor necrosis factor (TNF)-α (TNF), toll-like receptor 4 (TLR4), myeloid differentiation primary response gene (MyD)88 (MYD88), monocyte chemoattractant protein (MCP)1 (CCL2), peroxisome proliferator-activated receptor (PPAR)-α (PPARa) cluster of differentiation 36 (CD36), PPAR-γ (PPARg), PPAR-γ coactivator (PGC)-1α (PPARGC1a), ATP binding cassette subfamily G member 1 (ABCG1), and uncoupling protein 2 (UCP2), nuclear factor erythroid 2-related factor (NRF)2 (NFE2L2), NAD(P)H quinone dehydrogenase 1 (NQO1), heme oxygenase 1 (HMOX1) (Qiagen, Hilden, Germany), in a final volume of 50 μL. The relative amount of each studied mRNA was normalized to β-actin (ACTB) (Qiagen, Hilden, Germany) as housekeeping gene, and data were analyzed according to the 2−ΔΔCT method.
2.6. Statistical Analysis
All data shown are presented as mean value ± SEM. Intragender and intergender comparisons were made by two-way analysis of variance (ANOVA) followed by Bonferroni post hoc for multiple comparisons or Student t test, when appropriate. Differences among groups were considered significant at values of p <0.05. Analyses were performed using GraphPad Prism 9 (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. Gender Differences and LPS Effect on Biochemical Parameters and Inflammatory Mediators in Serum of WAG/Rij Rats
Intergender comparisons between male and female epileptic rats, exposed to LPS at PND3, showed a similar increase of lipid serum profile, such as elevated cholesterol and triglycerides (Figure 1A and B), as well as higher levels of hepatic transaminases ALT and AST, ALP, and LDH (Figure 1C-F). No difference was observed between female and male WAG without LPS challenge in Figure 1. Notably, we showed a marked susceptibility of male epileptic rats rather than females to LPS-induced systemic inflammation, as proven by the higher serum levels of twelve out of twenty-three pro- and anti-inflammatory cytokines analyzed by BioPlex assay (Figure 2A and B). Specifically, LPS-insulted males showed increased serum concentrations of pro-inflammatory mediators of innate immune activation (i.e. IL-1α, IL-12, IL-13) and T cell-derived cytokines, such as IFN-γ, IL-2, IL-4 and IL-5 and other markers produced in response to LPS challenge that regulate the mobilization of neutrophils and macrophages/monocytes against bacterial infection including M-CSF, and the beneficial IL-10 and IL-17A, suggesting the attempt to counteract the damage caused by LPS. No intragender differences in female groups were found (Figure 2A), while intergender comparisons between male and female groups without LPS evidenced higher concentrations of some inflammatory cytokines and mediators of innate and adaptative immunity in females than males, as reported in Supplementary Figure S2.
3.2. Hepatic Inflammation and Immune Response of Male and Female Epileptic Rats: Effect of an Early LPS Challenge
In the liver, we confirmed that male WAG/Rij rats were more prone to tissue inflammation and immune response to early LPS exposure compared with female animals. Indeed, we showed that LPS challenge markedly increased the transcription of IL-1β, COX-2 (Figure 3A, B), as well as that of TLR4 and MyD88 (Figure 3D, E) and the monocyte chemoattractant protein (MCP)1 (Figure 3F). Notably, no statistical intergender and intragender difference was found in the mRNAs of TNF-α (Figure 3C).
3.3. LPS-Driven Metabolic Alterations in Liver of Epileptic Rats: Sex-Related Differences
Conversely to LPS-induced inflammation, which was more pronounced in male epileptic animals, early endotoxin challenge induced a marked alteration in hepatic lipid metabolism of female WAG/Rij rats, as shown by a reduction of the mRNAs of the nuclear receptor PPAR-α (Figure 4A), mainly involved in the regulation of fatty acid oxidation (FAO), and the increased lipid transporter CD36 (Figure 4B), a key marker of steatosis, as well as PPAR-γ (Figure 4C). Moreover, LPS-insulted females showed a marked reduction of hepatic ABCG1 and UCP-2 transcription (Figure 4D and E), mitochondrial ATP-dependent transporter and liver-specific uncoupling protein, respectively. The mitochondrial damage was also suggested by the compensatory increase of PGC1-α (Figure 4F), a marker of mitochondrial biogenesis. Notably, intergender evaluations in unchallenged rats reveled a marked differences in hepatic lipid metabolism, as shown by the significant increase in PPAR-γ expression (Figure 4C) and the reduction of PGC1-α (Figure 4D) as well as the mitochondrial UCP2 (Figure 4F) in epileptic male rats rather than females.
3.4. Effect of LPS on Hepatic Mitochondrial Bioenergetics in Male and Female Epileptic Rats
LPS challenge induced the marked alteration of hepatic mitochondrial bioenergetics in females compared with males, as proven by the reduction of mitochondrial respiration in presence of succinate-rotenone and palmitoyl carnitine as substrates (Figure 5A,B), and the decrease of β-oxidation displayed by the reduced mitochondrial CPT activity (Figure 5C), as rate-limiting enzyme of FAO. A decrease in mitochondrial coupling was observed in female rats challenged with LPS (Figure 5D). Otherwise, in male epileptic rats, we noticed a strong release of hepatic mitochondrial H2O2 yield, while no differences were found in females (Figure 5E). A less prominent hepatic mitochondrial damage was induced by LPS infection in male rats, as showed by the slight decrease in mitochondrial function and no alteration in mitochondrial respiration, in presence of the specific substrates (Figure 5A,B) between WAG and WAG+ LPS groups, while fatty acid oxidative capability by CPT activity was increased in male rats exposed to LPS.
3.5. Sex-Related Susceptibility to Oxidative Stress of WAG/Rij Rats: Effect of LPS Injection
Finally, we demonstrated that LPS challenge induced oxidative damage in the liver of both genders increasing ROS production (Figure 6A). This LPS effect led to a gender-related activation of hepatic antioxidant defense, characterized by the increased mitochondrial SOD activity only in females (Figure 6B), and the altered hepatic glutathione redox status (GSH/GSSG ratio) in solely male WAG/Rij (Figure 6C). This latter finding was consistent with the increased H2O2 release, suggesting a remarkable oxidative damage in challenged male rats. Notably, LPS challenge differently modulated also the hepatic detoxifying system in female epileptic rats, increasing the gene expression of NRF-2 (Figure 6D) and the associated NQO1 (Figure 6E) and reducing HO-1 transcription (Figure 6F). No changes in detoxifying system were revealed in male animals, injected or not with LPS (Figure 6D- F), However, in LPS-challenged females the sustained antioxidant defense exerted by increased SOD activity and detoxifying factors cannot compensate the increase in free radical production (Figure 6A).
4. Discussion
Neonatal sepsis is a serious life-threatening condition and a major cause of morbidity and mortality. Immunity and metabolism play pivotal roles in the host response to infection and the severe metabolic demands of early life [23]; indeed, it has been hypothesized that the defense strategies differentially employed between newborns and adults can simply be attributed to differences in systemic energy supply and demand, revealing at the cellular level as differences in immune-metabolic activity [24].
The liver, as a highly immunocompetent organ, can activate a developmentally regulated innate immune response to LPS-induced sepsis [25,26], with the associated induction of pro-inflammatory mediators and pathways occurring only after the first month of life [26]. These findings support the long-term hepatic implications of early life exposure to systemic inflammatory stress. Indeed, gut-derived bacterial endotoxins, including LPS, can contribute to the pathogenesis of NAFLD and steatohepatitis by activating KCs [27]. Notably, a link between mortality in NAFLD-associated sepsis and hepatic mitochondrial and energetic metabolism dysfunction has been showed [13].
Here, we demonstrated the detrimental effect of early LPS challenge in inducing liver damage in young WAG/Rij epileptic rats, establishing the gender-based differences in hepatic and systemic inflammation and immune response, lipid dysmetabolism and associated mitochondrial oxidative damage. Contextually, these findings led us to characterize, for the first time, the gender metabolic profile of this useful strain regardless of early immunological damage.
WAG/Rij rats are a valid animal model of epileptic absences appearing from the first month of age (30-45 days) and share many electroencephalographic and behavioral features with epilepsy in humans, including a comparable response to different antiepileptic drugs and related side effects, including hepatotoxicity [17].
Recent studies have shown a vicious circle between liver damage and epilepsy; in detail, how seizures can complicate the course of liver disease, and in turn how liver damage can drastically reduce the therapeutic choice for epileptic patients. NAFLD is associated with a pro-inflammatory state and can induce peripheral and central inflammation causing neurotoxicity and the induction of seizures [12,28].
In our experimental conditions, male and female epileptic rats, early exposed to LPS, exhibited a compromised hepatic function, as clearly showed by increased serum transaminases AST and ALT, and LDH, as well as altered lipid profile. Otherwise, we evidenced a marked susceptibility of male WAG rats rather than females to systemic inflammation by LPS. In males LPS-challenge increased serum levels of pro-inflammatory cytokines of innate immune response and T cell-derived cytokines, and other mediators produced in response to LPS-challenge that regulate the release of neutrophils and macrophages/monocytes against bacterial infection. Moreover, an increase of beneficial IL-10 and IL-17A was shown, suggesting the effort by the organism to counteract the damage caused by LPS. These results agree with other findings in animals and humans, where neonatal males appear more susceptible than females to bacterial infections [14]. Male mice show an inappropriate inflammatory response to sepsis and produce significantly higher levels of pro-inflammatory cytokines than females following endotoxemia or sepsis [29,30,31]. Previous data reported that in animal model estrogen exerts a protective effect by maintaining adequate immune responses; conversely, ovariectomized rats are predisposed to sepsis and the addition of estradiol restored immune function [32,33]. Moreover, it has been showed that androgen receptor antagonism improves the compromised immune functions and reduces mortality for sepsis in both preclinical and clinical studies [34] In humans, the prevalence and severity of NAFLD are higher in men than in women during the reproductive age [35]. Indeed, after menopause, NAFLD occurs at a higher rate in women suggesting that estrogen may have a protective role. Generally, animal models of NAFLD resemble sex differences observed in patients with more severe steatosis and steatohepatitis, more pro-inflammatory/pro-fibrotic cytokines in males than females. In epileptic WAG/Rij rats, we confirmed the major predisposition of males than females to the inflammation and associated immune response primed by LPS at PND3 through the activation of COX-2, TLR4 pathway and macrophage/monocyte recruitment in the liver at PND45.
Notably, we found an intergender difference in epileptic rats without LPS about the serum concentrations of some inflammatory cytokines and mediators of innate and adaptative immunity which were more elevated in females than males. A different systemic or tissue control of inflammation by extra sexual hormones in unchallenged epileptic animals cannot be excluded.
It has been reported that the exposure to excess maternal fuels [36] or LPS [37] during fetal period causes the alteration of serum and hepatic lipid homeostasis, as well as liver morphology and mitochondrial health in adult mouse and rat offspring, respectively. These detrimental changes promote oxidative stress and excess triglyceride storage, along with immune dysfunction, that drive the hepatic damage progression from NAFLD to steatohepatitis in the adulthood [36].
Here, despite less evident alteration of serum lipid profile, early LPS infection led to the marked hepatic lipid dysmetabolism in female WAG/Rij rats rather than males, as shown by the opposite modulation of PPARs, differently involved in the regulation of lipid homeostasis, and increased lipid transporter CD36, a crucial marker of steatosis responsible for the influx of fatty acids into the hepatocytes. PPARs are considered metabolic sensors and therapeutic targets in different liver diseases [38]. Previous studies have recognized the implication of PPARγ in the development and maintenance of steatosis in the liver [39,40,41], since its downregulation in hepatocytes avoid cellular lipid accumulation [42,43]. Otherwise, PPARα, abundantly expressed in the liver and regulated during inflammation in a gender-specific manner [44], acts as a lipid sensor responding to the influx of fatty acids by inducing the transcription of other genes encoding for mitochondrial, peroxisomal, and microsomal oxidation systems [45,46,47].
Interestingly, in the liver of LPS-insulted females, we found a reduced expression of mitochondrial ATP-dependent transporter ABCG1 and uncoupling protein UCP-2, index of mitochondrial damage, also suggested by the compensatory increase of PGC1-α, as an attempt in preserving or ensuring an adequate mitochondrial biogenesis. These latter findings could explain the higher susceptibility of female rats to the detrimental effect of early LPS insult.
However, we proved that epileptic male rats without challenge showed an increase of PPARγ expression and a reduced PGC1α and UCP2 alteration of hepatic lipid metabolism than females.
Moreover, consistently with the alteration of lipid profile and metabolism, we strengthened a prominent hepatic mitochondrial damage of unchallenged male rats than females, which in turn revealed a major susceptibility to early immune infection in developing defects of mitochondrial bioenergetics in liver.
In female epileptic rats, the compromised mitochondrial respiration was evaluated in presence of specific substrates acting on different complexes of respiratory chain (succinate-rotenone and palmitoyl carnitine), and it was accompanied by the altered mitochondrial CPT activity, as rate-limiting enzyme of FAO. Notably, the two different gender-related mechanisms of LPS-induced liver injury (hepatic inflammation and lipid alterations) converge into oxidative damage in both sexes confirmed by increased hepatic ROS amount. Female epileptic rats, early exposed to LPS, exhibited a reduction in mitochondria efficiency indicated by a decreased degree of coupling. The electron transport and the ATP synthesis are processes that are closely coupled, but some of the energy generated by electron transport is uncoupled from ATP synthesis [48]. Instead, the reduction in mitochondrial efficiency allows the mitochondrial membrane potential to remain below the critical threshold for ROS production [49]. Indeed, the uncoupling is a major mechanism for the adjustment of the membrane potential to control mitochondrial ROS emission. With the observed mild uncoupling in female animals, the mitochondria can avoid the excessive supply of electrons/reducing equivalents in the respiratory complexes and minimize the probability of electron interaction with oxygen [48]. Consistently, the increased oxidative stress triggered a remarkable compensatory rise in hepatic antioxidant defense by SOD, which is the first line of defense from oxidative stress [50] and detoxifying enzymes only in female WAG/Rij animals. The different modulation of NRF2 pathway intermediates, such as NQO-1 and HO-1, suggest that the effect of an early immune challenge can negatively modulate the hepatic oxidative balance during epilepsy. Any improvement in the cellular redox state (GSH/GSSG) in the liver of female rats was shown.
It has known that LPS-induced sepsis and related inflammatory and stress conditions rises seizure susceptibility [51], considering the relevant role of oxidative stress and redox dysregulation in epileptic patients independently by antiepileptic pharmacological therapy[52]. Specifically, it has been found increased levels of oxidative biomarkers, including malondialdehyde, protein carbonylation and nitric oxide, in brain and peripheral tissues of both human patients and epileptic animals with any sex differences [52,53,54]. In the vicious relationship between epilepsy and oxidative damage, mitochondrial dysfunction plays a pivotal role, participating in the immunoinflammatory response [55,56]. It has been shown that many individuals with epilepsy exhibit concomitant mitochondrial disorders [57]. Furthermore, mitochondrial SOD2 knockdown in mice cause a remarkable rise in developing spontaneous motor seizures [58]. Moreover, the impairment of hepatic oxidative balance relieved mainly in male rats is confirmed by the altered GSH/GSSG ratio. The increase in GSH, usually occurs in response to oxidative stress, and a decrease in GSH can worsen disease [59]. Reduced GSH levels are also observed in several liver diseases [60], and in diseases associated with inflammation caused by microbial infections [61,62].
5. Conclusions
Summarizing, our data show, for the first time, a characterization of metabolic profile at systemic and hepatic level of WAG/Rij epileptic strain; contextually, we describe with a high translational potential that an early post-natal infection can predispose epileptic animals to develop or exacerbate hepatic disorders enhancing the different sex-related susceptibility. Female WAG/Rij have physiologically a higher serum levels of many inflammatory mediators and this difference was not evidenced in hepatic tissue and parameters, as well as lipid profile. Conversely, lipid metabolism was similar between female and male WAG without LPS challenge. However, LPS significantly impacts on lipid metabolism in female rats. Regarding mitochondria bioenergetics, a deep alteration in male rats, insulted or not with LPS, was shown, even if postnatal infection also reduced mitochondrial respiration and fatty acid oxidation in females. Despite the observed gender-related differences, our findings suggested that the effect of an early immune challenge can negatively modulate the hepatic oxidative balance during epilepsy in both genders, as demonstrated by the increase of ROS production.
Contemplating and then targeting the co-morbidity between liver damage and epilepsy can represent a fascinating health challenge also in the optimal therapeutic choice aimed at limiting the course of these pathologies, both characterized by a strong immune component.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Intergender evaluation of serum inflammatory/immune markers in WAG/Rij rats; Figure S2: Effects of LPS administration on (A) the number and (B) duration of characteristic SWDs in 3-month-old WAG/Rij rats.
Author Contributions
Conceptualization, C.P. and R.M.; methodology, S.M., G.T., C.P., G.M.R., M.P.M., R.M; validation, C.P., G.M.R., M.P.M., R.M.; formal analysis, A.L., M.P.M.; investigation, S.M., G.T, A.L., F.C., F.D.P., F.C., N.O., A.L., C.P.; data curation, S.M., G.T., A.L., R.C., C.P., M.P.M.; writing—original draft preparation, S.M., C.P., R.M.; writing—review and editing, C.P., R.M.; visualization, R.C., L.T., ; supervision, G.M.R., M.P.M., R.M.; project administration, R.M.; funding acquisition, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by funded by M.I.U.R.–PRIN: PROGETTI DI RICERCA DI RILEVANTE INTERESSE NAZIONALE – Bando 2017 Prot. 2017YZF7MA, Action line: South line, Unit responsible: Prof. Rosaria Meli.
Acknowledgments
We thank Mr Giovanni Esposito, Mr Angelo Russo, and Dr Antonio Baiano for animal care and technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
1. Supplementary Methods
1.1. Biochemical Evaluations of Serum Inflammatory/Immune Mediators
At PND45, blood was collected from all experimental groups. Then, serum was obtained by centrifugation at 2500 rpm at 4 ◦C for 12 min and stored at -80◦C for subsequent biochemical analyses. The concentration of twenty-three out of pro- and anti-inflammatory mediators, and factors of innate and adaptative immunity were obtained by a high sensitivity kit (Bio-Techne; R&D Systems, Inc., USA) using the Bio-Plex System and Luminex xMAP technology (Bio-Rad Laboratories, Inc., USA). Cytokine concentrations were derived by interpolating the measured fluorescence intensities to standard curves and correcting by the corresponding dilution factor employed to achieve the minimum volume for analysis. Bio-Plex Manager software was employed to calculate cytokine concentrations.
1.2. Electroencephalogram (EEG) Recordings of 3-Month-Old Male WAG/Rij Rats
Male WAG/Rij rat pups (n = 12) were randomly allocated into two subgroups; the first subgroup (n = 6) was intraperitoneally (i.p.) treated with LPS (from E. coli, serotype O55:B05, Sigma-Aldrich, MO) (1 mg/kg body weight) at three days of age (PND3). Vehicle rats received the same volume of sterile saline solution (100 μl). After the end of treatment, WAG/Rij rats were normally housed up to the age of ~3 months. Subsequently, LPS-treated and untreated WAG/Rij rats, under general anesthesia [mixture of tiletamine/zolazepam; 1:1; Zoletil 100®, 50 mg/kg i.p. (VIRBAC Srl, Milan, Italy)], were stereotaxically implanted with 3 cortical electrodes for EEG recordings attached to a 3-channel rat head-mount (8239-SE3; Pinnacle Technology, Stoke-on-Trent, UK) for seizure quantification, as previously described [1]. Quantification of epileptic seizures was based on the number and the duration of EEG spike-wave discharges (SWDs) according to well-established criteria. Briefly, the number and duration of SWDs (nSWDs and dSWDs) for each rat were summarized in 30-min intervals (epochs) and scored by visual inspection of the EEG recordings; all recordings were analyzed by two independent researchers that were blinded to the treatment [2].
1.3. Statistical Analysis
All statistical procedures were carried out using GraphPad Prism 10.0 software (GraphPad Software Inc., La Jolla, CA, USA). To assess LPS effects on absence seizures parameters, EEG recordings were subdivided into 30 min epochs, and both total and single duration and number of SWDs were assessed separately for every epoch. Such values were averaged, and data acquired were expressed as mean ± S.E.M. All tests used were two-sided, and p ≤ 0.05 was considered significant.
2. Supplementary Results
2.1. Intergender Differences on Inflammatory Mediators in Serum of WAG/Rij Rats
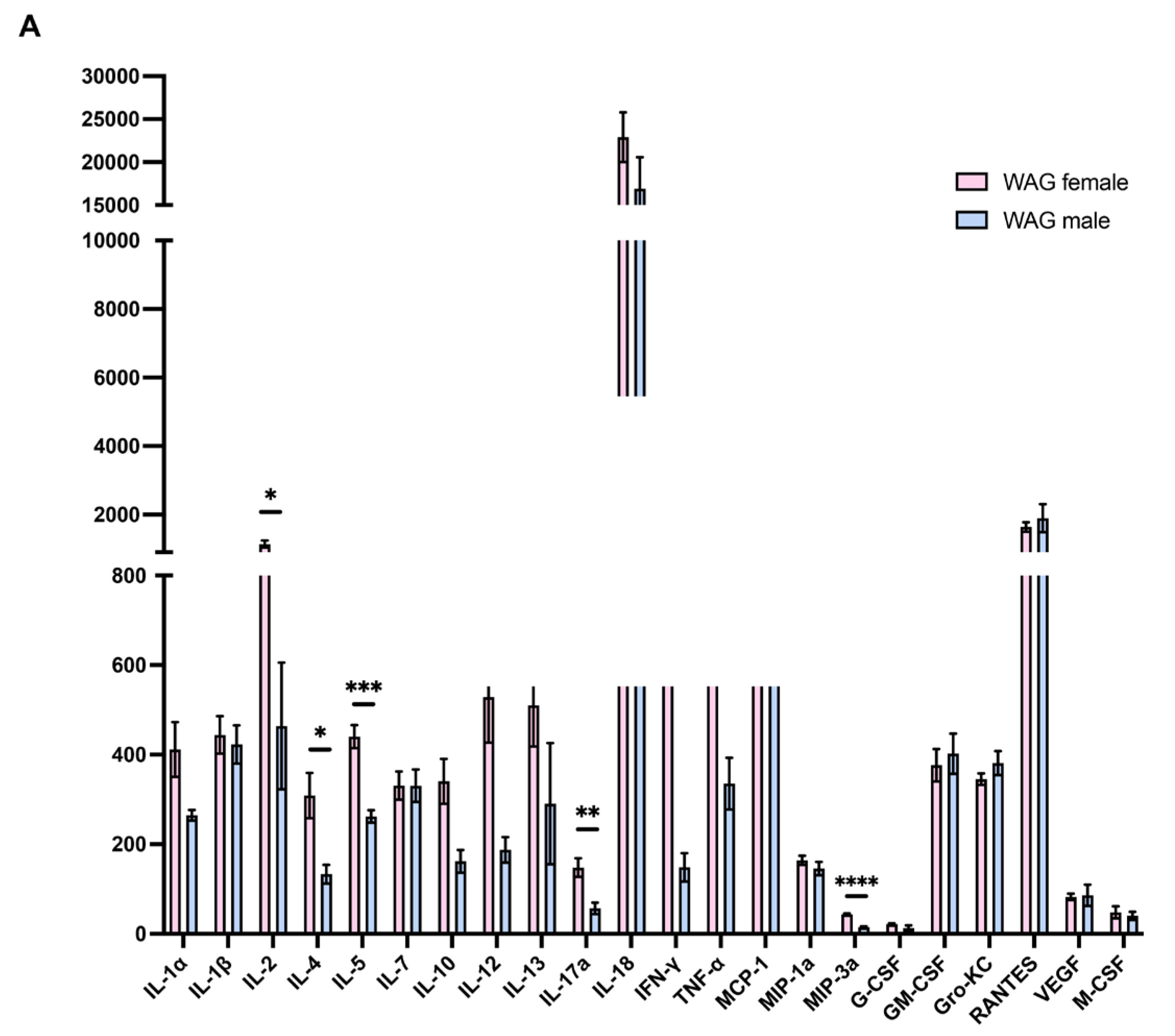
The intergender evaluation in epileptic rats without LPS by Bio-Plex assay revealed more elevated serum concentrations of T cell-derived cytokines, such as interleukin (IL)-2, IL-4, IL-5, and interferon (IFN)-γ, a mediator of innate immune activation such as IL-12, as well as macrophage inflammatory protein-3 (MIP3A) as a markers of macrophages/monocytes recruitment (Figure S1); these findings indicated a pronounced inflammatory state in female rats which could justify the less susceptibility to LPS challenge of females than males.
Figure S1.
Intergender evaluation of serum inflammatory/immune markers in WAG/Rij rats. Serum concentrations of inflammatory and immune markers were measured in male and female WAG/Rij rats by Bio-Plex System assay (n= 5-6 rats each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure S1.
Intergender evaluation of serum inflammatory/immune markers in WAG/Rij rats. Serum concentrations of inflammatory and immune markers were measured in male and female WAG/Rij rats by Bio-Plex System assay (n= 5-6 rats each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
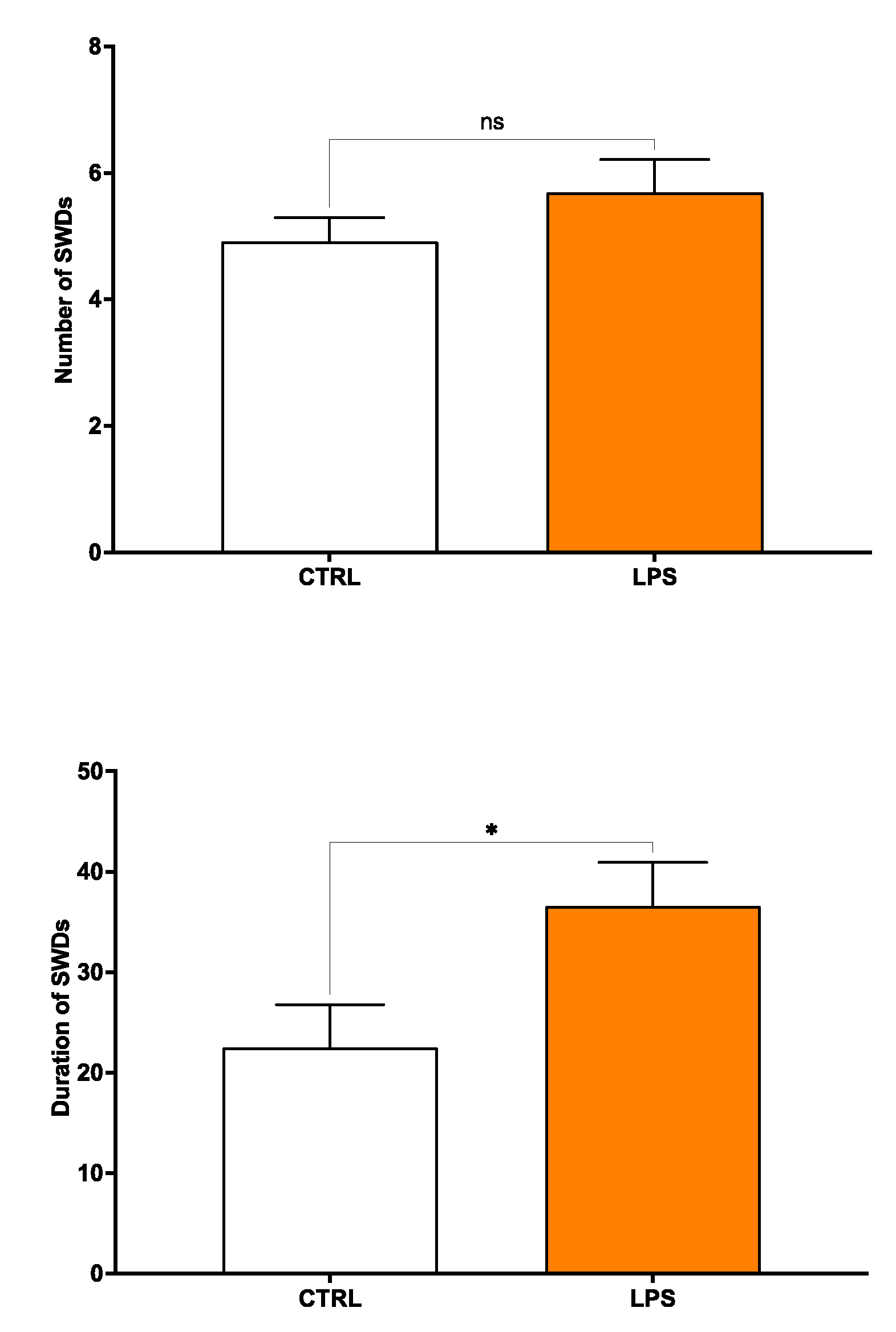
2.2. Early LPS Accelerates the Onset of Seizures in Young Adult Male WAG/Rij Rats
Preliminary results revealed no significant increase of nSWDs in LPS-challenged male WAG/Rij rats compared to age-matched control (vehicle) rats, while dSWDs were worsened by LPS injection in male epileptic rats. In detail, EEG recording analysis obtained from vehicle 3-month-old WAG/Rij rats (n = 6) showed a mean number of SWDs (nSWDs) of 4.91 ± 0.63, with a mean total duration (dSWDs) of 22.52 ± 6.06 s and a mean single duration (sSWD) of 4.24 ± 0.38 for a 30-min epoch. In contrast, acute treatment with LPS in WAG/Rij rats (n = 6) produced an increase of the mean nSWDs and dSWDs by ~ 14% (p = 0.25) and 39% (p = 0.04), respectively (Figure S2A-B). The mean sSWD value did not significantly differ between LPS-treated and untreated WAG/Rij rats, at this time point considered.
Figure S2.
Effects of LPS administration on (A) the number and (B) duration of characteristic SWDs in 3-month-old WAG/Rij rats. Data values are means ± SEM for every 30 min epoch (n = 6 each group). Data are presented as mean ± SEM. *p <0.05. CTRL = control; LPS = lipopolysaccharide; nSWDs = number of spike-wave discharges; dSWDs = duration of spike-wave discharges.
Figure S2.
Effects of LPS administration on (A) the number and (B) duration of characteristic SWDs in 3-month-old WAG/Rij rats. Data values are means ± SEM for every 30 min epoch (n = 6 each group). Data are presented as mean ± SEM. *p <0.05. CTRL = control; LPS = lipopolysaccharide; nSWDs = number of spike-wave discharges; dSWDs = duration of spike-wave discharges.
References
- Leo, V. Nesci, M. Tallarico, N. Amodio, E.M. Gallo Cantafio, G. De Sarro, A. Constanti, E. Russo, R. Citraro, IL-6 Receptor Blockade by Tocilizumab Has Anti-absence and Anti-epileptogenic Effects in the WAG/Rij Rat Model of Absence Epilepsy, Neurotherapeutics 17(4) (2020) 2004-2014.
- M. Tallarico, A. Leo, L. Guarnieri, M.C. Zito, C. De Caro, F. Nicoletti, E. Russo, A. Constanti, G. De Sarro, R. Citraro, N-acetylcysteine aggravates seizures while improving depressive-like and cognitive impairment comorbidities in the WAG/Rij rat model of absence epilepsy, Molecular Neurobiology 59(5) (2022) 2702-2714.
- Nakagaki, B.N.; Mafra, K.; de Carvalho, E.; Lopes, M.E.; Carvalho-Gontijo, R.; de Castro-Oliveira, H.M.; Campolina-Silva, G.H.; de Miranda, C.D.M.; Antunes, M.M.; Silva, A.C.C.; et al. Immune and metabolic shifts during neonatal development reprogram liver identity and function. J Hepatol 2018, 69, 1294–1307. [Google Scholar] [CrossRef] [PubMed]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat Rev Immunol 2017, 17, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Romics, L., Jr.; Frendl, G. Liver in sepsis and systemic inflammatory response syndrome. Clin Liver Dis 2002, 6, 1045–1066. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Jeong, W.I.; Tian, Z. Liver: An organ with predominant innate immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, M. Structural Modifications of Bacterial Lipopolysaccharide that Facilitate Gram-Negative Bacteria Evasion of Host Innate Immunity. Front Immunol 2013, 4, 109. [Google Scholar] [CrossRef]
- Song, H.; Zhang, X.; Zhai, R.; Liang, H.; Song, G.; Yuan, Y.; Xu, Y.; Yan, Y.; Qiu, L.; Sun, T. Metformin attenuated sepsis-associated liver injury and inflammatory response in aged mice. Bioengineered 2022, 13, 4598–4609. [Google Scholar] [CrossRef] [PubMed]
- Hastings, K.L.; Green, M.D.; Gao, B.; Ganey, P.E.; Roth, R.A.; Burleson, G.R. Beyond Metabolism: Role of the Immune System in Hepatic Toxicity. Int J Toxicol 2020, 39, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Luo, L.; Wang, Q.; Yan, S.; Lin, J.; Li, D.; Cao, B.; Mei, H.; Ying, B.; Bin, L.; et al. Maresin 1 attenuates mitochondrial dysfunction through the ALX/cAMP/ROS pathway in the cecal ligation and puncture mouse model and sepsis patients. Lab Invest 2018, 98, 715–733. [Google Scholar] [CrossRef] [PubMed]
- Markley, M.A.; Pierro, A.; Eaton, S. Hepatocyte mitochondrial metabolism is inhibited in neonatal rat endotoxaemia: Effects of glutamine. Clin Sci (Lond) 2002, 102, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Brealey, D.; Singer, M. Mitochondrial Dysfunction in Sepsis. Curr Infect Dis Rep 2003, 5, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Ferro, J.M.; Oliveira, S. Neurologic manifestations of gastrointestinal and liver diseases. Curr Neurol Neurosci Rep 2014, 14, 487. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, D.; Solmaz, V.; Taskiran, D.; Erbas, O. The association between seizure predisposition and inflammation in a rat model of fatty liver disease. Neurol Sci 2014, 35, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Zhang, J.; Cui, P.; Zhou, Y.; Liu, C.; Wu, X.; Ji, Y.; Wang, S.; Cheng, B.; Ye, H.; et al. TREM2 sustains macrophage-hepatocyte metabolic coordination in nonalcoholic fatty liver disease and sepsis. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Dias, S.P.; Brouwer, M.C.; van de Beek, D. Sex and Gender Differences in Bacterial Infections. Infect Immun 2022, 90, e0028322. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat Rev Immunol 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, K.; van Luijtelaar, G. The WAG/Rij strain: A genetic animal model of absence epilepsy with comorbidity of depression [corrected]. Prog Neuropsychopharmacol Biol Psychiatry 2011, 35, 854–876. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, C.; Lama, A.; Annunziata, C.; Cavaliere, G.; De Caro, C.; Citraro, R.; Russo, E.; Tallarico, M.; Iannone, M.; Ferrante, M.C.; et al. Butyrate prevents valproate-induced liver injury: In vitro and in vivo evidence. FASEB J 2020, 34, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Trinchese, G.; Cavaliere, G.; Cimmino, F.; Catapano, A.; Carta, G.; Pirozzi, C.; Murru, E.; Lama, A.; Meli, R.; Bergamo, P.; et al. Decreased Metabolic Flexibility in Skeletal Muscle of Rat Fed with a High-Fat Diet Is Recovered by Individual CLA Isomer Supplementation via Converging Protective Mechanisms. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Cairns, C.B.; Walther, J.; Harken, A.H.; Banerjee, A. Mitochondrial oxidative phosphorylation thermodynamic efficiencies reflect physiological organ roles. Am J Physiol 1998, 274, R1376–R1383. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Mattace Raso, G.; Cavaliere, G.; Trinchese, G.; De Filippo, C.; Aceto, S.; Prisco, M.; Pirozzi, C.; Di Guida, F.; Lama, A.; et al. Butyrate Regulates Liver Mitochondrial Function, Efficiency, and Dynamics in Insulin-Resistant Obese Mice. Diabetes 2017, 66, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal Biochem 1972, 48, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Bergamo, P.; Maurano, F.; Rossi, M. Phase 2 enzyme induction by conjugated linoleic acid improves lupus-associated oxidative stress. Free Radic Biol Med 2007, 43, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Harbeson, D.; Francis, F.; Bao, W.; Amenyogbe, N.A.; Kollmann, T.R. Energy Demands of Early Life Drive a Disease Tolerant Phenotype and Dictate Outcome in Neonatal Bacterial Sepsis. Front Immunol 2018, 9, 1918. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.G.; Angelidou, A.; Diray-Arce, J.; Smolen, K.K.; Lasky-Su, J.; De Curtis, M.; Levy, O. Immunometabolic approaches to prevent, detect, and treat neonatal sepsis. Pediatr Res 2020, 87, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Grayck, M.R.; McCarthy, W.C.; Solar, M.; Balasubramaniyan, N.; Zheng, L.; Orlicky, D.J.; Wright, C.J. Implications of neonatal absence of innate immune mediated NFkappaB/AP1 signaling in the murine liver. Pediatr Res 2024. [Google Scholar] [CrossRef] [PubMed]
- Zarate, M.A.; Nguyen, L.M.; De Dios, R.K.; Zheng, L.; Wright, C.J. Maturation of the Acute Hepatic TLR4/NF-kappaB Mediated Innate Immune Response Is p65 Dependent in Mice. Front Immunol 2020, 11, 1892. [Google Scholar] [CrossRef] [PubMed]
- Dembek, A.; Laggai, S.; Kessler, S.M.; Czepukojc, B.; Simon, Y.; Kiemer, A.K.; Hoppstadter, J. Hepatic interleukin-6 production is maintained during endotoxin tolerance and facilitates lipid accumulation. Immunobiology 2017, 222, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Fishel, M.A.; Watson, G.S.; Montine, T.J.; Wang, Q.; Green, P.S.; Kulstad, J.J.; Cook, D.G.; Peskind, E.R.; Baker, L.D.; Goldgaber, D.; et al. Hyperinsulinemia provokes synchronous increases in central inflammation and beta-amyloid in normal adults. Arch Neurol 2005, 62, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Aomatsu, M.; Kato, T.; Kasahara, E.; Kitagawa, S. Gender difference in tumor necrosis factor-alpha production in human neutrophils stimulated by lipopolysaccharide and interferon-gamma. Biochem Biophys Res Commun 2013, 441, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Oberholzer, A.; Keel, M.; Zellweger, R.; Steckholzer, U.; Trentz, O.; Ertel, W. Incidence of septic complications and multiple organ failure in severely injured patients is sex specific. J Trauma 2000, 48, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Frink, M.; Pape, H.C.; van Griensven, M.; Krettek, C.; Chaudry, I.H.; Hildebrand, F. Influence of sex and age on mods and cytokines after multiple injuries. Shock 2007, 27, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Knoferl, M.W.; Angele, M.K.; Schwacha, M.G.; Bland, K.I.; Chaudry, I.H. Preservation of splenic immune functions by female sex hormones after trauma-hemorrhage. Crit Care Med 2002, 30, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Knoferl, M.W.; Angele, M.K.; Diodato, M.D.; Schwacha, M.G.; Ayala, A.; Cioffi, W.G.; Bland, K.I.; Chaudry, I.H. Female sex hormones regulate macrophage function after trauma-hemorrhage and prevent increased death rate from subsequent sepsis. Ann Surg 2002, 235, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Angele, M.K.; Pratschke, S.; Hubbard, W.J.; Chaudry, I.H. Gender differences in sepsis: Cardiovascular and immunological aspects. Virulence 2014, 5, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, A.; Nascimbeni, F.; Ballestri, S.; Fairweather, D.; Win, S.; Than, T.A.; Abdelmalek, M.F.; Suzuki, A. Sex Differences in Nonalcoholic Fatty Liver Disease: State of the Art and Identification of Research Gaps. Hepatology 2019, 70, 1457–1469. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.R., 2nd; Friedman, J.E. Mitochondrial role in the neonatal predisposition to developing nonalcoholic fatty liver disease. J Clin Invest 2018, 128, 3692–3703. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wen, Y.; Li, J.; Zhang, H.; Liu, Y. Prenatal Lipopolysaccharide Exposure Promotes Dyslipidemia in the Male Offspring Rats. Front Physiol 2018, 9, 542. [Google Scholar] [CrossRef] [PubMed]
- Monroy-Ramirez, H.C.; Galicia-Moreno, M.; Sandoval-Rodriguez, A.; Meza-Rios, A.; Santos, A.; Armendariz-Borunda, J. PPARs as Metabolic Sensors and Therapeutic Targets in Liver Diseases. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Ohtake, T.; Motomura, W.; Takahashi, N.; Hosoki, Y.; Miyoshi, S.; Suzuki, Y.; Saito, H.; Kohgo, Y.; Okumura, T. Increased expression of PPARgamma in high fat diet-induced liver steatosis in mice. Biochem Biophys Res Commun 2005, 336, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Bedoucha, M.; Atzpodien, E.; Boelsterli, U.A. Diabetic KKAy mice exhibit increased hepatic PPARgamma1 gene expression and develop hepatic steatosis upon chronic treatment with antidiabetic thiazolidinediones. J Hepatol 2001, 35, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Rahimian, R.; Masih-Khan, E.; Lo, M.; van Breemen, C.; McManus, B.M.; Dube, G.P. Hepatic over-expression of peroxisome proliferator activated receptor gamma2 in the ob/ob mouse model of non-insulin dependent diabetes mellitus. Mol Cell Biochem 2001, 224, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Matsusue, K.; Haluzik, M.; Lambert, G.; Yim, S.H.; Gavrilova, O.; Ward, J.M.; Brewer, B., Jr.; Reitman, M.L.; Gonzalez, F.J. Liver-specific disruption of PPARgamma in leptin-deficient mice improves fatty liver but aggravates diabetic phenotypes. J Clin Invest 2003, 111, 737–747. [Google Scholar] [CrossRef]
- Matsui, J.; Terauchi, Y.; Kubota, N.; Takamoto, I.; Eto, K.; Yamashita, T.; Komeda, K.; Yamauchi, T.; Kamon, J.; Kita, S.; et al. Pioglitazone reduces islet triglyceride content and restores impaired glucose-stimulated insulin secretion in heterozygous peroxisome proliferator-activated receptor-gamma-deficient mice on a high-fat diet. Diabetes 2004, 53, 2844–2854. [Google Scholar] [CrossRef]
- Tai, E.S.; bin Ali, A.; Zhang, Q.; Loh, L.M.; Tan, C.E.; Retnam, L.; El Oakley, R.M.; Lim, S.K. Hepatic expression of PPARalpha, a molecular target of fibrates, is regulated during inflammation in a gender-specific manner. FEBS Lett 2003, 546, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Ohhira, M.; Motomura, W.; Fukuda, M.; Yoshizaki, T.; Takahashi, N.; Tanno, S.; Wakamiya, N.; Kohgo, Y.; Kumei, S.; Okumura, T. Lipopolysaccharide induces adipose differentiation-related protein expression and lipid accumulation in the liver through inhibition of fatty acid oxidation in mice. J Gastroenterol 2007, 42, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.S.; Reddy, J.K. PPARalpha in the pathogenesis of fatty liver disease. Hepatology 2004, 40, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.K.; Hashimoto, T. Peroxisomal beta-oxidation and peroxisome proliferator-activated receptor alpha: An adaptive metabolic system. Annu Rev Nutr 2001, 21, 193–230. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int J Mol Med 2019, 44, 3–15. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol Rev 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol 2024, 98, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- Sewal, R.K.; Modi, M.; Saikia, U.N.; Chakrabarti, A.; Medhi, B. Increase in seizure susceptibility in sepsis like condition explained by spiking cytokines and altered adhesion molecules level with impaired blood brain barrier integrity in experimental model of rats treated with lipopolysaccharides. Epilepsy Res 2017, 135, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Menon, B.; Ramalingam, K.; Kumar, R.V. Oxidative stress in patients with epilepsy is independent of antiepileptic drugs. Seizure 2012, 21, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Geronzi, U.; Lotti, F.; Grosso, S. Oxidative stress in epilepsy. Expert Rev Neurother 2018, 18, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Pearson-Smith, J.N.; Patel, M. Metabolic Dysfunction and Oxidative Stress in Epilepsy. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Sun, Y.; Li, L. Mitochondrial dysfunction in chronic neuroinflammatory diseases (Review). Int J Mol Med 2024, 53. [Google Scholar] [CrossRef] [PubMed]
- Fabisiak, T.; Patel, M. Crosstalk between neuroinflammation and oxidative stress in epilepsy. Front Cell Dev Biol 2022, 10, 976953. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S. Pathophysiology of mitochondrial disease causing epilepsy and status epilepticus. Epilepsy Behav 2015, 49, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.P.; Waldbaum, S.; Rowley, S.; Huang, T.T.; Day, B.J.; Patel, M. Mitochondrial oxidative stress and epilepsy in SOD2 deficient mice: Attenuation by a lipophilic metalloporphyrin. Neurobiol Dis 2012, 45, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Teskey, G.; Abrahem, R.; Cao, R.; Gyurjian, K.; Islamoglu, H.; Lucero, M.; Martinez, A.; Paredes, E.; Salaiz, O.; Robinson, B.; et al. Glutathione as a Marker for Human Disease. Adv Clin Chem 2018, 87, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Vairetti, M.; Di Pasqua, L.G.; Cagna, M.; Richelmi, P.; Ferrigno, A.; Berardo, C. Changes in Glutathione Content in Liver Diseases: An Update. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, S.C.; Zaretsky, M.D.; Dubs, J.G.; Roederer, M.; Anderson, M.; Green, A.; Mitra, D.; Watanabe, N.; Nakamura, H.; Tjioe, I.; et al. N-acetylcysteine replenishes glutathione in HIV infection. Eur J Clin Invest 2000, 30, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Mazari, A.M.A.; Zhang, L.; Ye, Z.W.; Zhang, J.; Tew, K.D.; Townsend, D.M. The Multifaceted Role of Glutathione S-Transferases in Health and Disease. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sex differences in serum biochemical and hepatic parameters in WAG/Rij rats following LPS challenge. Serum (A) cholesterol and (B) triglycerides, (C-D) transaminases, (E) ALP, and (F) LDH were measured in all experimental groups (n= 6-7 animals each group) by ELISA assay. Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure 1.
Sex differences in serum biochemical and hepatic parameters in WAG/Rij rats following LPS challenge. Serum (A) cholesterol and (B) triglycerides, (C-D) transaminases, (E) ALP, and (F) LDH were measured in all experimental groups (n= 6-7 animals each group) by ELISA assay. Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
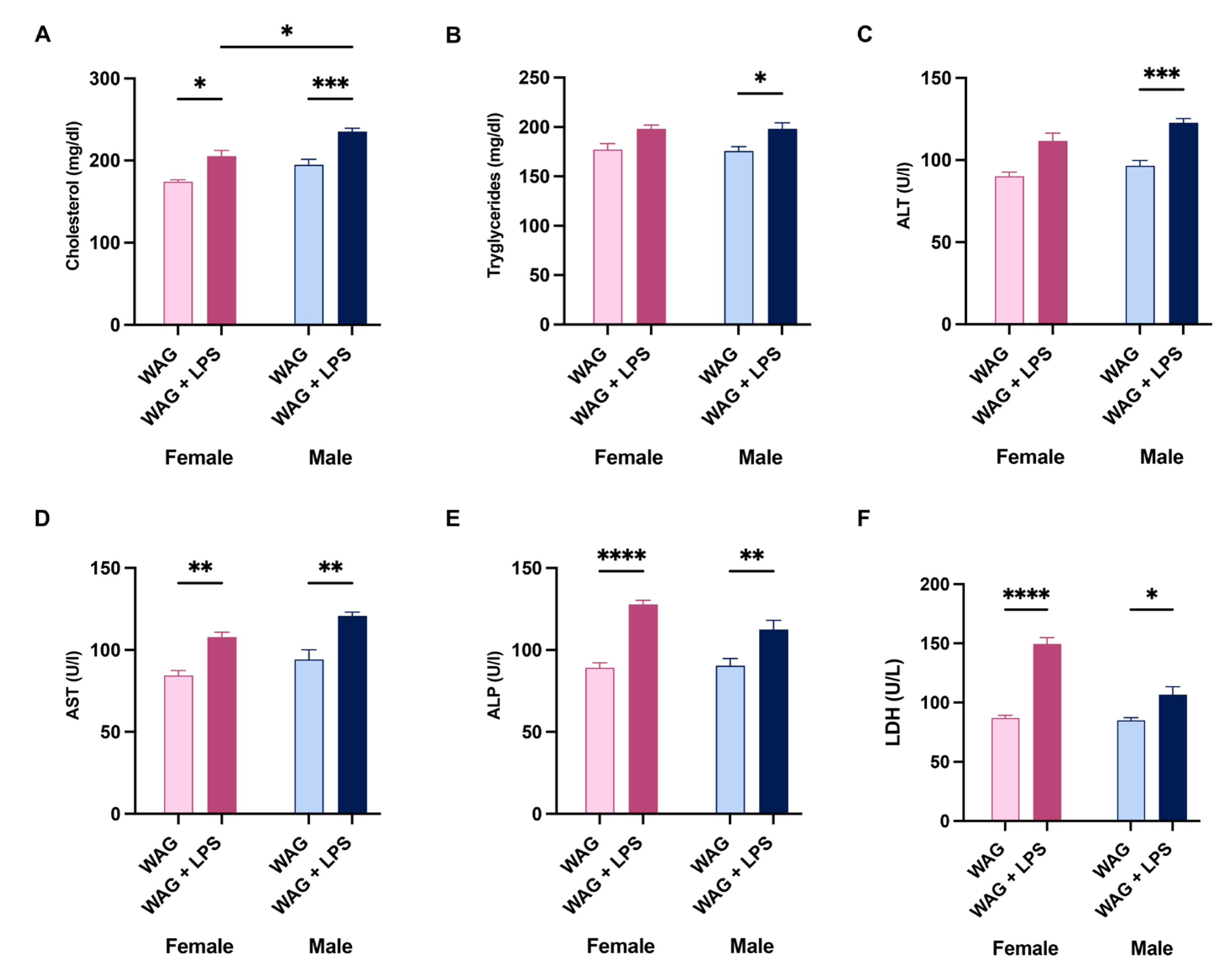
Figure 2.
Intragender modulation of inflammatory and immune markers in the serum of epileptic rats following postnatal LPS injection. The evaluation of systemic levels of inflammatory and innate/adaptative immunity mediators was performed by Bio-Plex assay in (A) female and (B) male WAG/Rij rats, challenged or not with LPS, respectively (6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0,0001.
Figure 2.
Intragender modulation of inflammatory and immune markers in the serum of epileptic rats following postnatal LPS injection. The evaluation of systemic levels of inflammatory and innate/adaptative immunity mediators was performed by Bio-Plex assay in (A) female and (B) male WAG/Rij rats, challenged or not with LPS, respectively (6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0,0001.
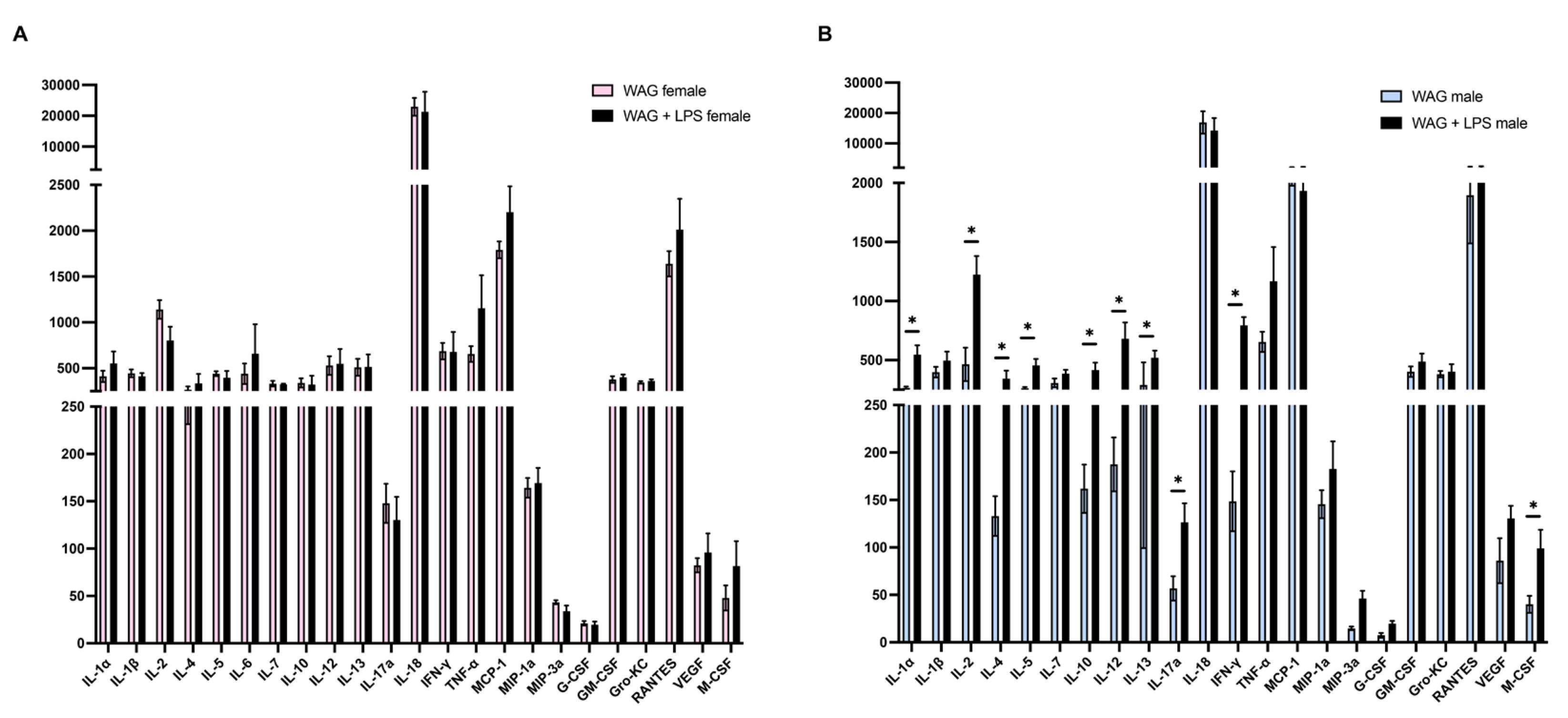
Figure 3.
Gender-related differences in hepatic inflammation and immune response of WAG/Rij rats: effect of neonatal endotoxin infection. mRNA expression of inflammatory (A) IL-1β, (B) COX-2, and (C) TNF-α, as well as (D) TLR4, (E) the coactivator MyD88, and (F) the chemoattractant MCP-1 was evaluated in the liver of all animals by Real-Time PCR (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure 3.
Gender-related differences in hepatic inflammation and immune response of WAG/Rij rats: effect of neonatal endotoxin infection. mRNA expression of inflammatory (A) IL-1β, (B) COX-2, and (C) TNF-α, as well as (D) TLR4, (E) the coactivator MyD88, and (F) the chemoattractant MCP-1 was evaluated in the liver of all animals by Real-Time PCR (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
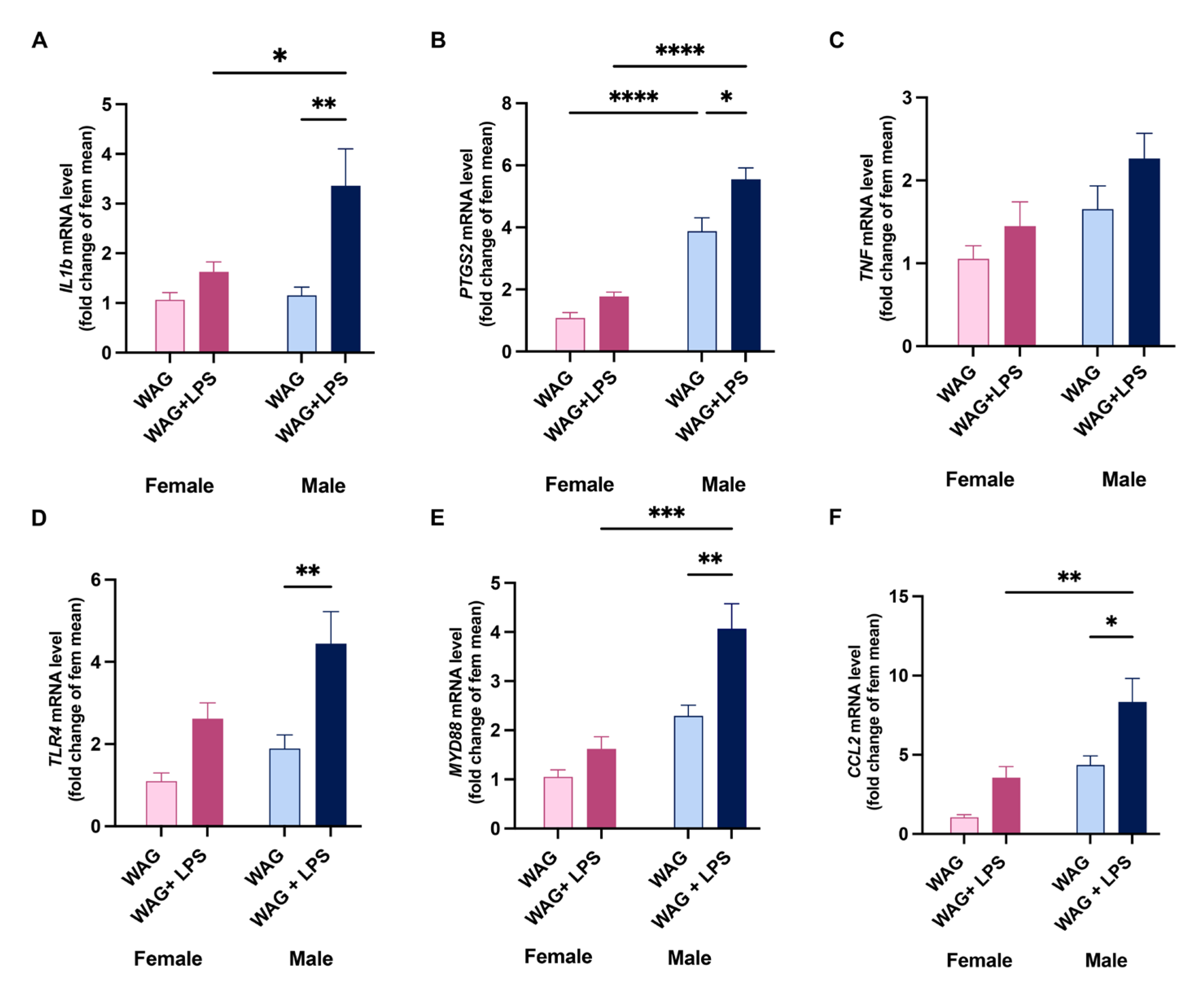
Figure 4.
Hepatic metabolic changes of male and female epileptic rats after or not early LPS challenge. Sex-related and LPS-induced differences in lipid metabolism and mitochondrial damage were assessed by Real-Time PCR analysis. Indeed, the transcription of (A) PPAR-α, and (B) CD36, as well as (C) PPAR-γ, (D) ABCG1, (E) UCP2, and (F) PGC1α was evaluated in all experimental groups (n=6-8 rats each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure 4.
Hepatic metabolic changes of male and female epileptic rats after or not early LPS challenge. Sex-related and LPS-induced differences in lipid metabolism and mitochondrial damage were assessed by Real-Time PCR analysis. Indeed, the transcription of (A) PPAR-α, and (B) CD36, as well as (C) PPAR-γ, (D) ABCG1, (E) UCP2, and (F) PGC1α was evaluated in all experimental groups (n=6-8 rats each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
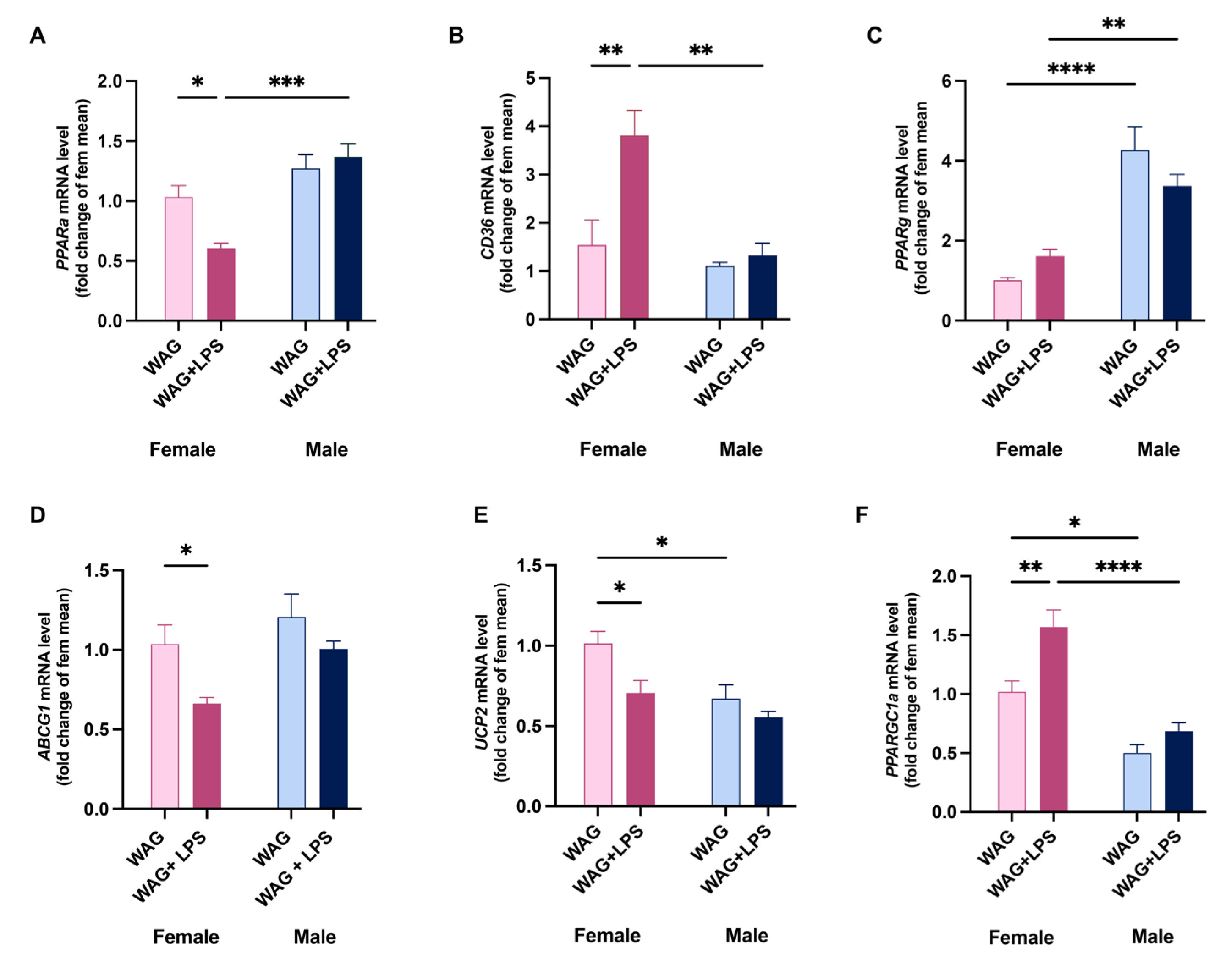
Figure 5.
Sex-based differences in hepatic mitochondrial bioenergetics of WAG/Rij rats following postnatal endotoxin injection. Mitochondrial respiration in presence or not of (A) succinate-rotenone and (B) palmitoyl carnitine as substrates was assessed in the liver of all animals by polarographic measurements (n=5-6 animals each group). (C) CPT activity, (D) the degree of coupling, and (E) H2O2 release into the isolated hepatic mitochondria of female and male WAG/Rij rats was measured by spectrofluorimetric analysis at PND45 (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure 5.
Sex-based differences in hepatic mitochondrial bioenergetics of WAG/Rij rats following postnatal endotoxin injection. Mitochondrial respiration in presence or not of (A) succinate-rotenone and (B) palmitoyl carnitine as substrates was assessed in the liver of all animals by polarographic measurements (n=5-6 animals each group). (C) CPT activity, (D) the degree of coupling, and (E) H2O2 release into the isolated hepatic mitochondria of female and male WAG/Rij rats was measured by spectrofluorimetric analysis at PND45 (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
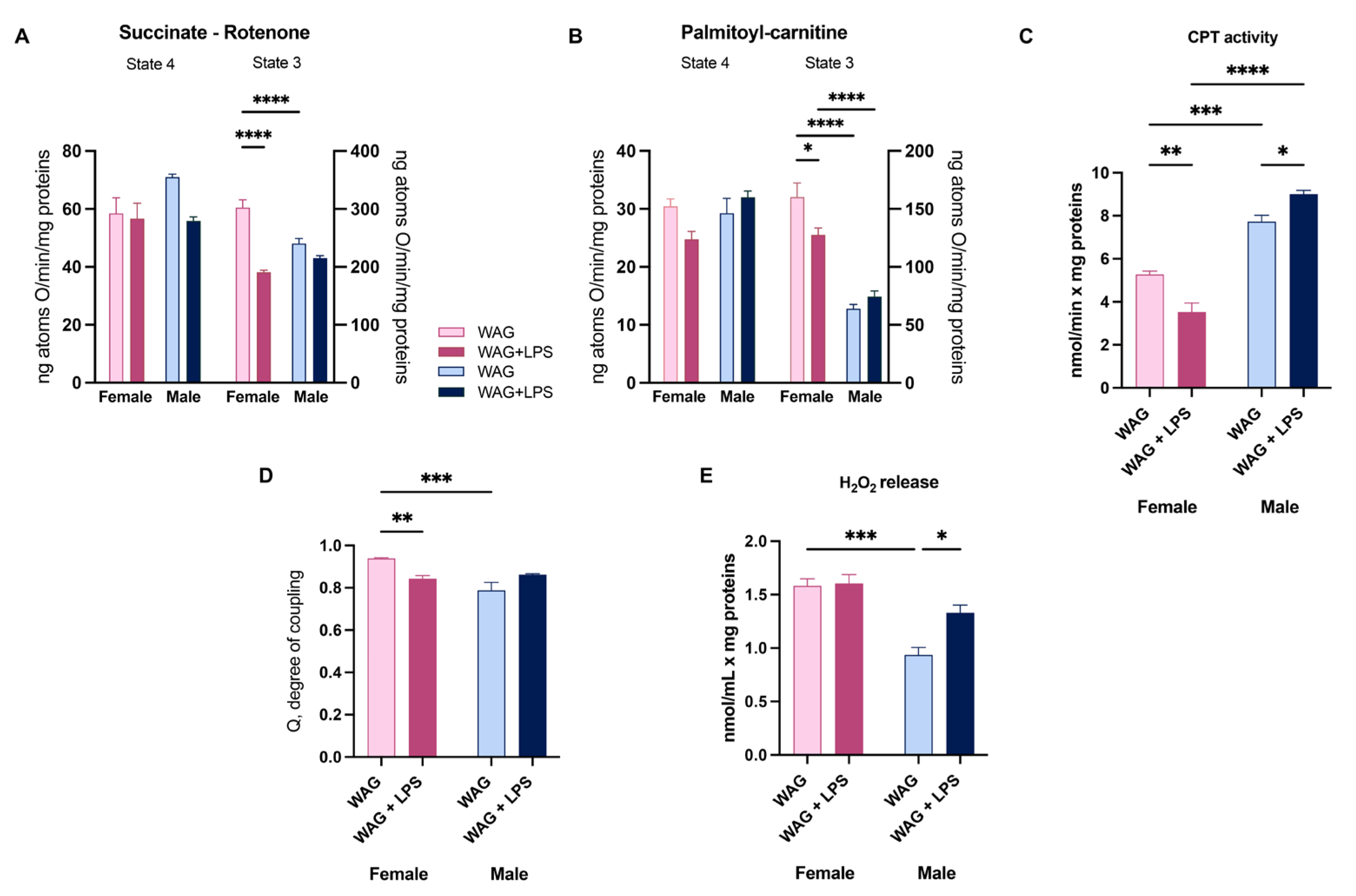
Figure 6.
Hepatic oxidative stress and anti-oxidant defenses in epileptic rats: gender differences and effect of early LPS challenge. (A) ROS production, as well as (B) mitochondrial SOD activity were spectrofluorimetrically measured in the liver of female and male WAG/Rij rats, challenged or not with LPS (n= 6-8 animals each group). Moreover, (C) hepatic glutathione redox status (GSH/GSSG ratio) was calculated in all experimental groups (n=5-6 animals each group). The mRNA transcription of detoxifying factors (D) NRF-2, (E) NQO1, and (F) HO-1 was assessed by Real-Time PCR at PND45 (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Figure 6.
Hepatic oxidative stress and anti-oxidant defenses in epileptic rats: gender differences and effect of early LPS challenge. (A) ROS production, as well as (B) mitochondrial SOD activity were spectrofluorimetrically measured in the liver of female and male WAG/Rij rats, challenged or not with LPS (n= 6-8 animals each group). Moreover, (C) hepatic glutathione redox status (GSH/GSSG ratio) was calculated in all experimental groups (n=5-6 animals each group). The mRNA transcription of detoxifying factors (D) NRF-2, (E) NQO1, and (F) HO-1 was assessed by Real-Time PCR at PND45 (n=6-7 animals each group). Data are presented as mean ± SEM. *p <0.05, **p <0.01, ***p <0.001, and ****p <0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



