
Submitted:
07 December 2023
Posted:
08 December 2023
You are already at the latest version
Abstract
An event of increasing interest during host-pathogen interactions is the differentiation of patrolling monocytes (MOs) and the polarization of naive MOs into macrophage subsets (MФs). Therapeutic strategies aimed at modulating this event are under investigation. This review focuses on the mechanisms of induction, development, and profile of MФs polarized toward classically proinflammatory (M1) or alternatively anti-inflammatory (M2) phenotypes in response to bac-teria, fungi, parasites, and viruses. It highlights nuclear (PPARγ), cytoplasmic, and cell surface receptors (pattern recognition receptors/PPRs), microenvironmental mediators, and immune signaling. MФs polarize into phenotypes: M1 MФs, activated by IFN-γ, pathogen-associated mo-lecular patterns (PAMPs, e.g. LPS)/DAMPs and membrane-bound PPRs ligands (TLRs/CLRs lig-ands); or M2 MФs (a, b, c and d), induced by IL-4, -10 and -13, antigen-antibody complexes, and helminth PAMPs. The kinetics of M1 and M2 polarization evolve in a pathogen-specific manner, with or without canonicity, and can vary widely. Ultimately, this can result in varying degrees of host protection or more severe disease outcome. While the host is driving an effective MФs po-larization, pathogens are attempting to shift it to increase pathogenicity. The latter results from a bias toward an ineffective M1 or M2 response due to microbial virulence factors, culminating in pathogen evasion of host defenses. Plasticity of M1/M2 phenotypes is also ensured by cellu-lar/genomic reprogramming. Because modulation of this event can occur at multiple points, providing a therapeutic/vaccine design target to boost microbicidal (M1) or tissue repair (M2) response during the inflammation-to-resolution transition; new insights and emerging perspec-tives may have clinical implications.
Keywords:
Macrophage
; polarization
; receptor
; bacteria
; virus
; fungi
; parasites
1. Introduction
Infectious diseases remain a serious public health problem worldwide, responsible for a significant proportion of morbidity and mortality. They also have a significant impact on populations in terms of disability-adjusted life years (DALYs)/years lived with disability (YLDs), sequelae and quality of life [1,2,3,4].
During the innate immune response, the action of monocytes (MOs) and MOs differentiated into macrophages (MФs) has been reported as an important weapon of the human host against various pathogens. It has been extensively studied in the context of infections caused by viruses, protozoa, bacteria, and fungi [5,6,7,8]. These cells are involved in wound healing, the transition between innate and acquired immunity, and the maintenance of tissue homeostasis. They also fight potentially pathogenic microorganisms and regulate the inflammatory process associated with infectious diseases [9,10,11].
The classification of MФs, first reported in 2000, is based on the polarization of CD4+ T-helper (Th) lymphocytes and the corresponding Th1 and Th2 responses proposed in 1986. In recent literature, the M1 (formerly known as classically activated MФs) and M2 (alternatively activated MФs) phenotypes have been described as subpopulations, subtypes, or subsets of MФs. This model is based on MФs ability from C57BL/6J to produce nitric oxide (•NO) (M1 phenotype) versus MФs ability from BALB/c mice (M2 phenotype) [10,11,12]. However, as of 2014, a more cautious view has shown that the dichotomous model appears minimalistic given the different transition and continuity phenotypes that exist in a population of MФs during the immune response, regardless of whether a canonical dynamic is present or not. This demonstrates the complexity of the broad spectrum of MФs, which may occur depending on the complex and multifactorial chemical microenvironment that permeates inflammatory and microbial stimuli in each tissue [11,12,13,14]. It is temporally delimited by the kinetics and specificity of a particular innate immune response and its importance in the transition to the acquired response. The current evidence has led to the proposal of a multi-polarization model of MФs. This has already been demonstrated for a more heterogeneous population of CD4+ Th cells (Th1, Th2, Th17) [11,14,15,16,17,18].
Microbial stimuli such as pathogen-associated molecular patterns (PAMPs) and/or microenvironmental signals, immune mediators, trigger mechanisms through specialized immune pathways and drive MФs polarization. Among these stimuli/inducers produced by the host during infection, damage-associated molecular patterns (DAMPs, such as ↑[ATP], succinate, HMGB1, cardiolipin, n-formyl peptides, TFAM, mitochondrial DNA and histones) are highlighted, Resolution-associated molecular patterns (RAMPs, such as HSP 10 and 27) and specialized pro-resolution mediators (SPMs, including resolvins, protectins, molesins, lipoxins, and other lipid mediators) are also involved in MФ polarization; as these, in addition to PAMPs and other microbial stimuli, are also recognized by pattern recognition receptors (PRRs, such as toll-like receptors, TLRs, TLR4 involved in recognition of lipopolysaccharide (LPS) or HMGB1 (DAMPs), or RAMPs; mitochondrial DNA recognized by TLR9), P2X7 [e. e.g. recognition of DAMPs (ATP) and PAMPs], nuclear receptors [NRs, such as peroxisome proliferator-activated receptor gamma (PPARγ) and liver X receptors (LXRs)] and other cell surface receptors (CSRs) in MOs/MФs, providing targets for immunophenotyping [8]. In MOs/MФs, membrane-bound PRRs include TLRs (1, 2, 3, 4, ….9) and C-type lectin receptors (CLRs). Cytoplasmic PRRs are represented by NOD-like receptors (NLRs) and RIG-I-like receptors (RLRs) [6].
In this way, the broad polarization profile of MФs can be demonstrated by the specific structural features and markers of these cells, such as the cluster of differentiation (CD), as well as by the distinct functional phenotypes. A highly microbicidal/proinflammatory phenotype has been characterized for M1 MФs or anti-inflammatory for M2 MФs [8]. Thus, M1 MФs are microbicidal, while M2 MФs are pro-resolving and can mitigate tissue damage by promoting tissue repair and healing (i.e., they can also fight some pathogens, e.g., helminths). However, during polarization and repolarization (reprogramming), both profiles (M1 and M2) work together in the transition from inflammation to resolution of inflammation/infection [8].
In this review, we discuss the stimuli for inducing, activating, and identifying the phenotypes of MФs populations during the innate immune response to medically relevant microorganisms and clinical implications. We will highlight the importance of PAMPs, PRRs, microenvironmental signals, and other immune mediators involved in the immune pathway as well as microbial virulence factors against MФs immune response.
2. MOs, MФs and polarization: an overview
Leukocytes are a diverse group of blood cells. They mediate the body's immune response. Among leukocytes, ~10% are MOs [19]. These hematopoietic cells of the mononuclear myeloid lineage are estimated to have an average residence time in the blood stream of 1 to 3 days. In addition to being considered as antigen presenting cells (APCs) that recognize pathogens and present their derivatives in lymph nodes near when they leave the bloodstream, MOs give rise to specific tissue MФs (Table 1). These MФs perform important constitutive functions in the human organism that are controlled by different pathophysiological states, location, and specific signals from the microenvironment, with a lifespan that can vary from a few days to months or years (Table 1) [20]. The tissue MФs themselves also serve as APCs carrying information about the context of the tissue infection to the nearest lymph node, with the aim of reinforcing the immune response [21]. Identified more than 100 years ago by Ilya Metchnikoff, whose description of phagocytosis earned him a Nobel Prize in Medicine in 1908, MФs stand out for recognizing and neutralizing aggressive stimuli (phagocytosis and microbicidal activity), producing immunomodulatory substances (such as those that afford the adaptive immune response) and preserving the integrity of different tissues in the face of an exacerbated response (through resolution/repair/healing) [6,22,23].

Table 1.
Tissue distribution of macrophages (MФs) in humans.
| Location* | Resident cells | Main microorganisms (stimuli/tropism) that activate the response |
| CNS (brain) | Microglia** |
Toxoplasma gondii, Schistosoma spp., Cryptococcus neoformans, virus, and bacteria |
| Bones | Osteoclasts*** | Anaerobic microorganisms, S. aureus |
| Heart/vessels | Resident cardiac MФs | Trypanosoma cruzi, Streptococcus spp., Candida albicans, Staphylooccus aureus |
| Liver | Kupffer cells | Plasmodium spp., Trypanosoma spp., Schistosoma spp., HAV, HBV, HCV, S. aureus |
| Lungs | Alveolar MФs | Mycobacterium tuberculosis (Mtb), Aspergillus fumigatus, Pseudomonas aeruginosa, S. aureus, C. neoformans, Streptococcus spp., SARS-CoV e SARS-CoV-2, other viruses |
| Adipose tissue**** | Adipose tissue MФs | Brucella spp., parasites, SARS-CoV-2 |
| Connective tissue | Histiocytes | Pathogens in general |
| Intestine/ Peritoneum | MФs associated with the intestine/Peritoneal MФs | Enterobacteriacae, some viruses, parasites in general; immunotolerance to commensals |
| Kidneys | Mesenchymal cells | Virus and bacteria |
| Spleen | Red pulp MФs (RPMs) | Plasmodium spp., parasites, and microbes of blood origin |
| Skin/Epidermis | Langerhans cells/ Dendritic cells (DCs-MOs) |
Staphylooccus spp., Mycobacterium leprae, Leishmania spp., Paracoccidioides brasiliensis, Candida spp. |
*Resident macrophages are more abundant in the CNS and epidermis and less abundant in the intestine and dermis. In the latter two sites, circulating monocyte-derived macrophages (MФs-MOs) predominate. In the other sites, there is a balance between resident macrophages and MФs-MOs [19]. In all cases, resident MФs are present in both normal and pathological conditions. The type and frequency of MФs may vary depending on the microenvironment and the type and chronology of the aggression. **According to Ransohoff [23], the minimalist model of M1 and M2 microglial nomenclature should be avoided. It neglects the more complex issues that exist for the spectrum of MФs subpopulations in the CNS, which takes into account the implications of the pathophysiological state and functional machinery of these cells. ****In the context of viral and bacterial infections, these cells can be activated. This includes acting as a reservoir for microorganisms.
In a context that goes far beyond the old models of classically activated and alternatively activated MФs, the nuances of activation and development of polarization of specific and functional subpopulations of these cells can be didactically divided into several stages. In more advanced stages, reprogramming can occur, making the population profile of MФs in a given microenvironment heterogeneous and favoring all the processes in which these cells participate. Differentiation of circulating mature MOs into tissue MФs is the first stage. MOs are exposed to various stimuli designed to prime these cells. This is done by stimuli that may not be strong enough to generate a specific polarization, but that will make the cells more susceptible to other specific stimuli that they will encounter in the infectious focus and/or target anatomical site (second phase). In a third phase, the MФs reach a mature functional profile in response to a specific stimulus, and those MФs that survive the defense task enter a deactivation profile, also called resolution of the inflammatory/infectious focus. During the resolution phase, those M1 MФs that survived the infectious-inflammatory focus can be reprogrammed to the M2 profile, i.e. that of repair and healing, in addition to a subpopulation of M2 that is polarized (mainly by programming circulating MOs attracted by chemokines). Another hypothesis is that tissue MФs are at rest (M0) under physiological conditions or even in the early stages of pathological processes. In this context, functional reprogramming occurs. Thus, when appropriately primed and stimulated, these cells can be classically activated (M1), alternatively activated (M2a), or deactivated (M2c) [24,25].
While the phenotypic sub-populations of M1 MФs are avidly related to microbicidal activity, in the so-called "common host response against pathogens", mainly in the acute phases initiating and sustaining inflammation, presenting high production of reactive oxygen (ROS) and nitrogen (RNS) species, combined with a high expression and activity of inducible nitric oxide synthase (iNOS), the MФ2 sub-populations are related to the resolution of the infectious/inflammatory process and tissue repair and, in the specific response to different stimuli from parasites, fungi and intracellular bacteria and viruses (Table 2) that infect vital organs or tissues, presenting high gene expression and functional activity of arginase (Arg) and considered immunomodulatory [21]. Therefore, congruent with the orchestrated immune response in humans, M1 MФs are related to the Th1-type response pattern and NK cells (cellular response), while M2 MФs subpopulations are reported to have a Th2-type response pattern and interact with B lymphocytes (stimulating the humoral response), mast cells and basophils (mediating allergic processes), as well as responding via type 2 inflammation in response to certain microorganisms [10,11,26,27,28].
There are no unanimous reports in the literature on a single or a few membrane markers for evaluating the differentiation of MOs into MФ1 and MФ2 subpopulations, and contextual evaluation (also reporting the species from which the MФ originates), which involves evaluating a set of membrane markers, the biological context from which the MOs or MФs originate (e.g. inflammatory focus, presence of cancer, regenerated tissue, among others), the functional activity of these cells (oxidative state, microbicidal activity, arginase/iNOS ratio, among others) and gene expression analysis can guide the interpretation of a particular MФ subtype in a cell population. Even so, the membrane markers CD80, CD86, MHC class II (also present in lower density on the MФ2 cell surface) are more closely related to the MФ1 subtype and CD23, CD163 and CD200R to MФ2 [10,18].
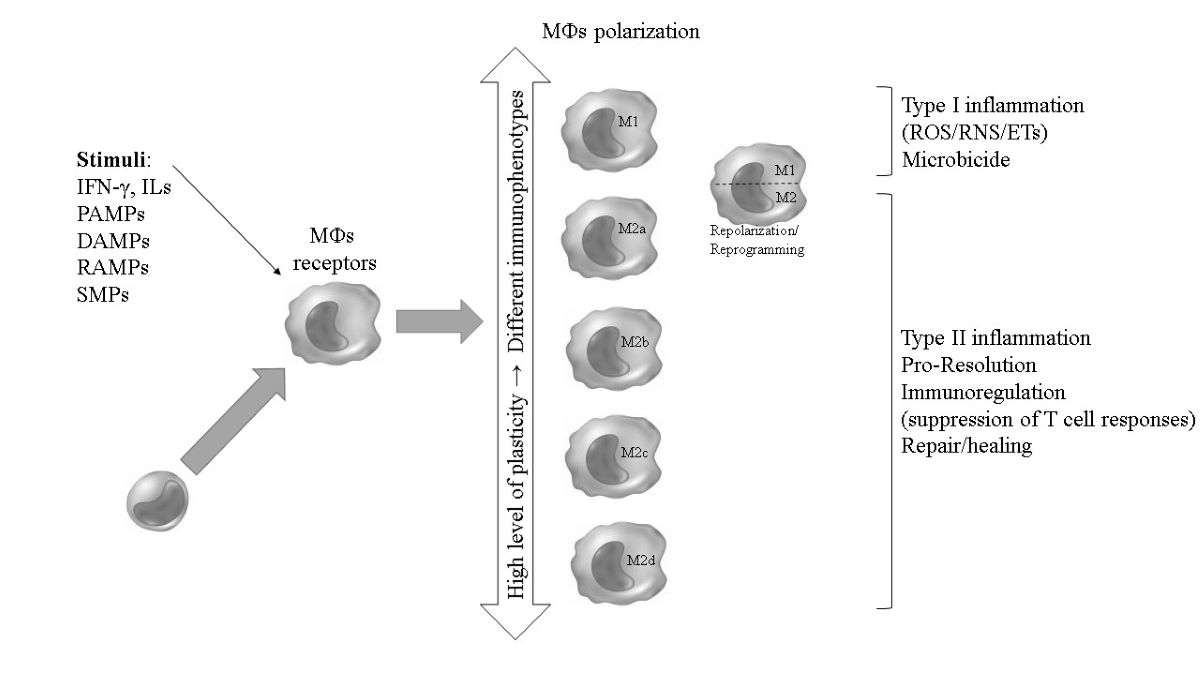
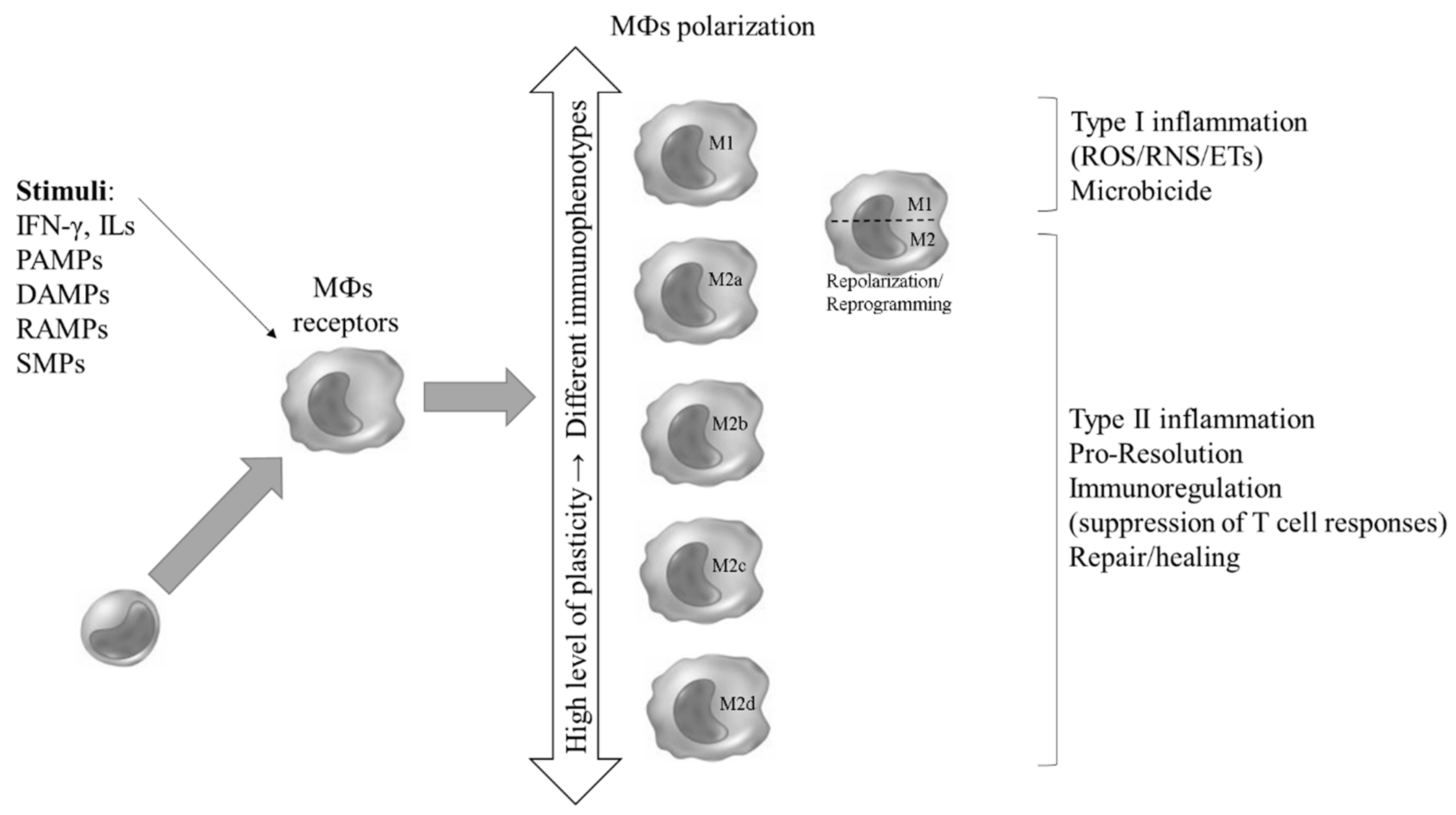
It should be noted that, due to these multiple characteristics performed by MФs, with various cellular and molecular components responsible for the primary recognition of a pathogen (innate response), activation of microbicidal mechanisms and signaling for an orchestrated specific (acquired) and memory response, in addition to the resolution of the infectious and inflammatory focus and tissue healing and growth, it has been suggested that, in vivo, there is not just one dichotomous profile of these cells, but rather different polarized sub-populations (M1; M2a, b, c and d, Mhem and MOx; Table 3 and Figure 1) that coexist at a given site of infection, and that, temporally, some sub-populations may stand out numerically. This fact has been confirmed in studies exploring different biological contexts using specific experimental models, in which there is a great heterogeneity of MФs subtypes, but with the possibility of a certain subtype prevailing. Thus, the paradigm of the coexistence of sub-populations in vivo is also the result of a temporal issue in the response to tissue aggression, since a prolonged and excessive polarization of the M1 type that occurs in the common host response to pathogens, leads to a consequent profile of M2 programming aimed at the production of anti-inflammatory mediators and resolution of the infectious focus [29].
2.1. Stimuli for the activation of polarization
Irrespective of whether the immune response involved is T-cell dependent or independent, as well as whether it involves other cell types (such as the T lymphocyte-independent B lymphocyte response), the scope of this review is to emphasize in detail and exclusively the role of macrophages in polarization; thus ignoring the potential coexistence of other responses, such as when the antigen is protein/peptide and activates T cells to direct and amplify the response, or when the antigen is lipid/carbohydrate and activates a response via B cells with a predominantly humoral profile. It should be noted that circulating MOs and tissue MФ, such as professional phagocytes, respond immediately to microbial stimuli. Thus, in a manner that is not necessarily canonical [30], PAMPs of protein/peptide origin tend to generate a response that involves T cells, which may at some point modulate the targeting and amplification of the polarized response via a particular MФ profile. Microorganisms with non-protein PAMPs, i.e. carbohydrates or lipids, usually induce a Th2/M2 skewed response in addition to the primary phagocytic response. This response can also involve a T lymphocyte-independent B lymphocyte response, i.e. in response to potentially pathogenic fungi (Candida albicans and its glycans), parasites (containing glycans) and bacteria (Escherichia coli containing LPS) (PAMPs and stimuli summarized in Table 2) [30,31]. In general, after recognition of microbial PAMPs, the immune response is processed and directed through the production of cytokines. Naive MOs and MФs are also primed with cytokines and/or proinflammatory stimuli of the Th1 profile, including interferon (IFN)-γ produced by such cells (recognized by IFN-γR1 and 2), TNF-α, IL-1β, 6 and 12; polarizing to M1 MФs, while those cells primed with cytokines IL-4, 10 and 13 are polarizing to M2 MФs [30,31].
Table 2.
Stimuli and receptors involved in macrophage (MФs) polarization.
| PAMP/DAMP RAMP/SPM |
Origin | Macrophage receptors/location | Macrophage action |
|---|---|---|---|
| LPS, spike, formyl peptides, flagellin, high mannose, chitin, β-glucans, ss/dsRNA, cpg dna | Lipids, proteins, carbohydrates, and nucleic acids from microorganisms | PRRs: TRLs (2, 3, 4, 5, 7, 8, 9, 11) and CLRs (membrane-bound) and NLRs/RLRs (cytoplasmic); Dectin-1 | Proinflammatory PAMPs. |
| DNA, RNA, IL-1A/B, histones, HSPS, uric acid, oxidized phospholipids, decorin, fibronectin | Nucleus, cytoplasm, plasma membrane and extracellular matrix of the dying cell | PRRs (TLRs 2, 4, 7, 8, and 9), RAGE, NLRP, CDs, P2X7, ↑IL1RLI, IL1RLII | DAMPs with inflammation-inducing activities. |
| HSP10, HSP27 | Dying cell, tissue-resident cells | TLR4, CD36, MSR, MERTK, PTGERE | Pro-resolution RAMPs. |
| lipoxins, resolvins, protectins, maresins | Efferocytes, tissue resident mesenchymal stromal cells | LGR, GPRs (18, 32, and 37), ALX, ERV | SPMs with resolution-inducing activities. |
| HMGB1, ↑[ATP], IL-33, PGE2, annexin1 | Dying cell |
TLR 2, 4, 5, RAGE, TREM-1, EP2, 4, ST2, FPR2 | DAMPs and RAMPs acting in the transition from inflammation to resolution |
PAMP: pathogen-associated molecular patterns; DAMP: damage-associated molecular patterns; RAMP: resolution-associated molecular patterns; SPM: specialized pro-resolution mediators. Dectin recognizes β-glucans from pathogenic fungi. Reference: revised in [8].
Thus, the induction of MФs polarization is guided primarily by the specificity of the stimulus that induces it (Table 3), unless during the course of such a programmed response there are factors that modify it, such as those presented in the repertoire of microorganisms [11], or by the host itself in order to control an exacerbated response and/or accelerate tissue resolution and repair (reprograming polarization). In that way, PAMPs can induce a specific polarization, acting directly through interaction with PRRs in T and B lymphocytes, as well as MOs and/or MФs [28,32]. Among the PRRs, the TLRs, mainly TLR4, have been the most described. PAMPs (such as LPS) can act indirectly on polarization, modulating the cytokine/chemokine profile and/or inducing IFN-γ which will subsequently interact with MФs.
Furthermore, the ability of MФs to recognize and respond to a wide variety of stimuli in specific microenvironments is the result of a wide range of receptors present on the surface of these cells, such as those above mentioned for the recognition of microorganisms (including PRRs), for coagulation factors, for components of the extracellular matrix, transport proteins, growth and differentiation factors and for cytokines [33]. Also, after the MФs are stimulated, cytoplasmic mediators are activated, such as nuclear factor (NF)-κB, and gene expression is activated, defining the phenotypic profile of the MФ, as will be discussed below.
Table 3.
Spectral model of macrophage (MФs) polarization in humans and their associated profiles.
| Phenotype | Activation/ Stimulus |
Markers | Immune signaling and molecules/functionally: Transcriptional profile and cytokine/chemokine production | Profile functional of the phenotype | |
|---|---|---|---|---|---|
| M0 | ---- | ---- | ---- | ---- | |
| M1 | IFN-γ;a PAMPs/LPS; GM-CSF; other TLR ligands |
CDs 68, 80, 86; MHC II; |
↑iNOS; TLR2, TLR4; ILs-1β, 6, 8, 12, and 23; TNF-α, IFN-γ; CXCL 8, 9, 10, 11, and 16; CCL 2, 3, and 5; |
Microbicide Type I inflammation Inflammasome ↑Oxidative burst (ROS/RNS) ↑ETs M1/Th 1,17 responses |
|
| M2a | ILs-4 and -13; Fungi/ Helminths |
CDs 23, 163 e 200R | ↑ARG1; IGF1; DecoyR; ILs-1r, and10; TGF-β; IL-CCL17, 22, and 24; |
Resolution of infection Killing and encapsulation of parasites Allergy M2/Th2 responses Type II inflammation |
|
| M2b | IC+TLR/ (Ac-Ag); IL-1R IL-1β LPS |
CD86 MHC II |
↑eNOS; CCL1; ILs- 1, 6, 10 , and 16; TNF-α |
Resolution of infection/inflammation Immunoregulation M2/Th2 responses |
|
| M2c | IL- 10; TGF-β; Glucocorticoids | CD163 TLR1, R8 |
ILs-1β and 10; TGF-β; CCR2; MMP9; ↑ARG1 |
Immunoregulation (suppression of T cell responses) ↑Repair/healing |
|
| M2d | IL-5; LIF; Adenosine |
VEGF | ILs-10 and 12; TNF-α; TGF-β; CCL5; CXCL10 and 16 | ---- | |
| M4 | ---- | CDs 86 and 206 | TNFα; CCL18 and 20 | ---- | |
| Mhem | ---- | CD 163 | HO-1; IL-10 | ---- | |
| MOx | ---- | ---- | HO-1, SD-1, TR-reductase | ---- | |
a The different types of IFN can have different functions and induce different subsets of MФs during polarization.
Figure 1.
Graphical Abstract. Macrophages (MФs) (re)polarization in response to different stimuli and the associated functional profiles of their immunophenotypes. .
Figure 1.
Graphical Abstract. Macrophages (MФs) (re)polarization in response to different stimuli and the associated functional profiles of their immunophenotypes. .
2.2. Profile of polarized MФs: development and ramifications in programming and reprogramming sub-populations
After responding to a given stimulus, MФs will mount a cellular response that begins in the cytoplasm and extends to the activation of specific genes that guide the production of proteins and mediators that influence cellular metabolism, the signaling of other cells and the response profile to the aggressive stimulus. Programming is understood as the specific cellular machinery as a whole [stimulus uptake receptors (such as PRRs), second messengers (such as kinases), gene expression and metabolism regulation sensors (such as NF-қβ, HIF-1, AMPK and mTOR)] that coordinately direct certain functional properties [proinflammatory (M1 MФs) or anti-inflammatory (M2 MФs) profile] of polarized MФs [26,34]. In 1938, Kempner was the first to link cell metabolism and inflammation [28]. Therefore, programming encompasses the entire context of the activity of MOs and polarized MФs and can be initiated through the priming of circulating MOs by cytokines and extending to the tissues. It can also begin in the tissues and reprogramming, an event that reverts to the functional characteristics achieved by a certain subpopulation of MФs, is similar to programming in that it has transcriptional and metabolic characteristics. As it involves a broad context, programming and reprogramming will be discussed in different topics.
2.3. Metabolic programming and transcriptional profile during the differentiation of MOs into MФs and polarization
2.3.1. Differential gene expression and metabolism
Polarization programming and reprogramming is a dynamic and rapid process that involves signaling networks at the transductional, transcriptional and translational levels. In a context that involves an intricate cytoplasmic network for the transduction of the signal captured by MФs membrane receptors, and nuclear, governed by the specificity of the stimulus, transcriptional factors and post-transcriptional regulators are highlighted. Monocyte and MФ NRs are ligand-activated transcriptions factors and key players to control polarization in infectious diseases. IFN regulatory factor (IRF), signal transducers and activators of transcription (STAT) and suppressor of cytokine signaling (SOCS) are some of these. The IRF/STAT pathway triggered by IFN-γ or TLR (induction by microorganisms) converges on the activation of pathways involving NF-қB/PI3K, triggering the transcription of genes related to the M1 program (as examples, the TNF-α, COX2, CCL5 NOX2 and iNOS genes, and for the cytokines IL-1 and IL-6). On the other hand, MФs activated with IL-4 or IL-13 trigger STAT6 and downstream elements, such as PPARγ and δ, which inhibit the transcription of genes of the M1 program (for example, by trans-repression of NF-қB) and trigger those of the M2 program (such as ARG1), which is potentially anti-inflammatory [28,35]. A broad transcriptional profile was reported by Martinez et al [26], revealing the intense transcription of genes related to the proinflammatory profile in M1 MФs and anti-inflammatory in M2 MФs.
Regarding the metabolic machinery, the metabolism of proteins and amino acids, lipids, carbohydrates and even minerals and the redox status markedly differentiate the subpopulations of MФs [28]. Among the changes in metabolic programs, those in amino acid and carbohydrate metabolism have been described as important markers of the different MФs subpopulations. Thus, the enzymes IDO1 and IDO2 and TDO, which catalyze the oxidative catabolism of the amino acid tryptophan (Trp), are important markers of amino acid metabolism and M1 and M2 functional profiles. The actions of these enzymes imply a microbicidal capacity by restricting the bioavailability of Trp, which is essential for the growth of a wide variety of microorganisms (viruses, bacteria, and parasites), therefore, in the infectious focus, high expressions delimit the sub-population phenotypic profile of M1 MФs [28]. A similar strategy has been described for iron (Fe) metabolism in the infection microenvironment, where M1 MФs repress the expression of ferroportin (an Fe transporter) and CD163 (an Fe-rich hemoglobin scavenger receptor) and induce ferritin (which favors the intracellular sequestration and storage of Fe in MФs), consequently sequestering (competing for Fe) this essential element for the microorganism and also controlling the redox context in which Fe participates [28].
The enzymes Arg1 and iNOS also play important roles as metabolic and functional indicators of MФs, related to competition for the metabolism of the amino acid arginine. Thus, when MФ expresses more iNOS to the detriment of Arg1 (MФ1 sub-population), L-arginine will be used preferentially for the synthesis of •NO or urea (using NADPH and O2), thus orchestrating the intricate microbicidal capacity of these cells. On the other hand, when MФ expresses more Arg1 to the detriment of iNOS (M2 MФ sub-population), the production of ornithine or L-citrulline will be favored, ornithine being a precursor of proline which, along with ornithine itself and other polyamines formed, are essential for collagen biosynthesis and tissue growth and fibrosis (healing MФs), thus guaranteeing the functionality of M2 MФ during tissue repair and remodeling [10,11,26,28,34,36].
As for energy metabolism, the control of hypoxia, which is common in the microenvironment of lesions due to infection and/or inflammation and high metabolic requirements, is also in the repertoire of MФs [28]. The stabilized hypoxia-inducible factor-1 (HIF-1) induces the expression of hundreds of genes that regulate angiogenesis, metabolism, and survival, favoring adaptation to low O2 pressures and the microbicidal action of MФs. In this same context, the energy sensors: AMP-activated kinase (AMPK, controls ATP synthesis) and the target of rapamycin in mammals (mTOR, adapts to protein synthesis); are capable of adapting the metabolism of MФs in infectious-inflammatory microenvironments to the demand required by the adversities of these contexts and also to avoid the energetic benefit of the microorganism. A decrease in AMPK activity is congruent with a proinflammatory population phenotype (M1 MФs), while an increase is anti-inflammatory (M2 MФs) [28]. Thus, the metabolism of carbohydrates is strongly influenced by the context of polarization (also involving mitochondrial dynamics), as well as that of lipids, which suffers metabolic consequences during β-oxidation and, above all, presents specific membrane lipid components due to the signaling of the inflammatory process [25,37]. As an example, the β-oxidation pathway in M2 MФs has been described as functional in the activity against some parasites [34].
2.3.2. Cytokine and chemokine profile during polarization
Cytokines such as interleukins (ILs) and TNF-α, and homologous cytokines such as chemokines (chemotactic factors with CC or CXnC structural sequences) during the innate immune response mediate various functions of the immune system, also serving as a communication mechanism between cells and allowing the transition between innate and acquired immunity. The effects of these mediators as well as their transcriptional profile are strongly distinct in different phenotypic subpopulations of MФs [26]. Thus, the profile of proinflammatory cytokines and chemokines is related to M1-type polarization and anti-inflammatory to M2 [28]. In this context, after being polarized, MФs also produce the same cytokines that stimulated them, amplifying and converging the response pattern [11]. As examples, ILs and TNF-α are the main mediators of the acute inflammatory response, and therefore correlate with the population phenotypes of MФ M1, which consequently stimulate Th1 lymphocytes [27].
2.3.3. Reprogramming of the transcriptional and metabolic system and repolarization
The question has been raised that, both for circulating MOs with an initial commitment to a certain functional program, when they reach the target tissue/inflammatory focus, as well as for tissue MФs, there may be a transcriptional and functional reprogramming of the population in question with a change in the circulation or microenvironment in terms of the cytokine profile (such as ILs and IFN-γ) and other mediators [34,37]. This implies that the same population of M1 MФs can acquire characteristics of M2 and vice versa, depending on the change in microenvironment and timing, a fact that highlights programming as a reversible process, as long as the MФs are not compromised by apoptosis, ETosis or another form of death [28]. This phenomenon has been highlighted mainly in sub-populations of MФs influenced by the microenvironment and its alterations, as an adaptation response to the new context, also observed in infectious/inflammatory processes and their transitions, and/or in a context where the microorganism makes use of virulence factors and alters polarization, thus changing the type of response and prolonging the disease, as will be discussed later [28,29]. In addition, the human organism itself can reprogram polarization, with the aim of avoiding irreversible damage to the tissue/organ during an intense/prolonged cytotoxic response, or based on a response via RAMP/SPM/DAMP-sensing receptors that are expressed by both MФs, M1 and M2 [34,37,38].
2.4. Polarization of tissue MФs
The classic definition of tissue MФs as cells capable of performing an inflammatory and antimicrobial response led to the concept of two stages: a basal or dormant stage (M0) and an activated stage [25]. However, recent studies have shown that there are other metabolic and functional stages of these cells, which go beyond a simple model of the dormant-activated duality. These studies reveal a gradual spectrum [M1, M2 (from a to d subtypes), M4, Mhem, and MOx] between predominantly dormant (non-activated) populations (Table 3), passing through a state in which predominantly proinflammatory (activated) populations prevail, followed by a predominance of anti-inflammatory or pro-regenerative (deactivated) populations, in a temporal context that depends on host factors, the stimulus that induced the response and the persistence of the response [39]. Furthermore, the priming of MOs still in circulation can predetermine a specific polarization of MФs in their targeting of specific infected tissues.
2.5. Functional profile of polarized MФs
The functional profile, a result of the context of the microenvironment and kinetics of the response, is considered one of the main arguments for polarization, as will be seen below.
2.5.1. Phenotypic subpopulations of M1 MФs: phagocytosis and microbicidal activity (intra- and extracellular killing)
2.5.1.1. Microbicidal activity
Microbicidal activity has been described as one of the main characteristics of M1 MФs. Different elements are highlighted during this response (phagocytosis, oxidative burst and oxidative burst-associated other microbicidal activities, as well as non-oxidative killing), as presented below.
2.5.1.1.1. Phagocytosis
Phagocytosis following the recognition of foreign bodies in the organism is the initial milestone in the proinflammatory and microbicidal response. It is an event that allows the aggressive stimulus to be internalized and, in a tautochronous manner, signals an appropriate immune response. Although classically related to the M1 MФs profile, this process can also be carried out by M2 MФs, in those microbial infections in which this functional profile is considered immunoprotective, but M2 MФs are less predisposed to carrying out phagocytosis [11,28].
2.5.1.1.2. The NADPH oxidase complex: microbicidal profile of oxidants, activation of enzymes with antimicrobial activity and formation of extracellular traps, as well as signaling in the immune system
Phagocytes exert two different microbicidal mechanisms: oxidative and non-oxidative. As discussed so far, the profile of M1 MФs is considered proinflammatory and highly microbicidal, in this context, one of the most important mechanisms of the inflammatory and antimicrobial response performed by these cells is the production of oxidants during the respiratory/oxidative burst [40]. During the oxidative microbicidal response, the NADPH oxidase complex (Nox2 system in phagocytes) generates O2•- through the reduction of molecular oxygen (O2) and, through local action, this weaker oxidant generates other ROS, such as hydrogen peroxide (H2O2) and, in addition, through the action of the myeloperoxidase (MPO), reactive intermediates are formed, such as the hydroxyl radical (HO•), hypochlorous acid (HOCl) and molecular singlet oxygen (1Δg O2). These last three oxidants are more potent antimicrobials, also known as highly reactive oxygen species (hROS). Furthermore, with activation of the iNOS enzyme, RNS such as •NO are formed, with •NO reacting rapidly with O2•-, giving rise to ONOO-, a vigorous antimicrobial agent [41,42]. In fact, the direct microbicidal effect of oxidants, causing oxidative damage to the microbial cell, can contribute to the intracellular killing of pathogens. At the same time, signaling for the activation of enzymes with antimicrobial activity, based on the redox context provided during the oxidative burst, is pointed out as a synergistic microbicidal effect to the direct action of oxidants. Although the oxidative burst of MOs and M1 MФs is less intense than that produced by neutrophils, this mechanism performed by MOs/M1 MФs is still fully effective in combating pathogenic microorganisms. Furthermore, reinforcing the importance of the microbicidal profile of oxidants produced by M1 MФs in effectively fighting infections, primary (chronic granulomatous disease, CGD) or secondary deficiencies of the NADPH oxidase complex may predispose humans to primo-infections and recurrent infections by Staphylococcus aureus, Gram negative bacilli, fungi of the genera Aspergillus and Candida, and even more uncommon potentially pathogenic microorganisms such as Burkholderia cepacia, Chromobacterium violaceum, Nocardia and more invasive strains of Serratia marcescens [40,43].
In addition, representing a recently discovered mechanism in the field of Immunology, first reported in the late 1990s [44] but further elucidated and named by Brinkmann et al [45], the oxidants generated through the oxidative burst also activate the so-called extracellular traps (ETs), highlighting yet another of the synergistic microbicidal mechanisms performed by phagocytes [46,47]. It is now known that ETs are produced by neutrophils (neutrophil extracellular traps, NET), mast cells (mast cells extracellular traps, MCET), eosinophils (eosinophil extracellular traps, EET) and MOs/MФ, with the latter cells generating the so-called MO/MФ extracellular traps (METs), first reported by Chow et al [48]. Since then, several studies have shown that MOs, MФs in general and tissue MФs of different species can generate ETs when exposed to a wide variety of stimuli, including chemical (e.g. PMA) and microbial (including their PAMPs/derivatives such as LPS, the tripeptide fMLP and the leukocidin Panton-Valentine) [42,49,50,51,52,53,54,55]. ETs are made up of decondensed chromatin complexed with protein granules with antimicrobial action [from azurophilic (elastase and myeloperoxidase), specific (lactoferrin) and tertiary (cathepsin and gelatinase) granules]. The formation of these structures seems to respond specifically to different pathogen/chemical-specific stimuli and the robustness of this response also varies with the type of phagocyte in question, with ETs being able to capture microorganisms in the extracellular space and, in certain cases, promote their extracellular killing, whether or not accompanied by the programmed death of the MOs/MФs, in which case it is referred to as ETosis [56,57].
Therefore, during the microbicidal response, the intense production of ETs can be considered a marker of the M1 MФs profile. Furthermore, as recently demonstrated, taking into account the classic context in which neutrophils are the first cells to respond to a tissue aggression, the presence of the antimicrobial peptide LL37 in NETs or other ETs, can recruit MФs-MOs to the inflammatory site and promote an M1-type polarization [58]. On the other hand, Guimarães-Costa et al [59] reported that NETs formed against Leishmania spp. can prevent the differentiation of MФs-MOs into M1 DCs-MOs, altering the functional profile of the cells toward an anti-inflammatory one and consequently decreasing the leishimanicidal activity (probably in order to decrease the cytotoxic response). Also, given this context of activation and the complex patterns of pathogen-specific immune response, including those in which M2 MФs are protective against some pathogens, and considering that ETs can be induced by different routes, including those independent of ROS [60], it is not ruled out that ETs formed by M2 MФs may contribute to the immune response and prevent the spread of the pathogen.
Evidence also points to the importance of the NADPH oxidase complex and consequent production of oxidants, shown in the profile of M1 MФs during the innate response and the transition to an acquired Th1-type response [61]. As attested by Neil et al. [62], the use of Tempol, a potent antioxidant, decreased oxidant levels in an experimental model of multiple sclerosis, which also altered the transition from an acquired response to a Th2 profile, contrary to what was expected, which would be a Th1 profile, in which case there is an immunomodulatory rather than immunosuppressive mechanism exerted by the nitroxide. Furthermore, as evaluated by Tillack et al. [63], the ETs produced can directly establish a link between innate and acquired immunity through priming. The programming of M1/M2 and Th1/Th2 patterns is also cited as being induced and dependent on the oxidants generated during the oxidative burst [64].
2.5.1.1.3. Induced nitric oxide synthase (iNOS)
Considered one of the main mediators generated by populations of M1 MФs, the high gene expression and consequent activity of iNOS are congruent with the microbicidal profile of these cells. iNOS forms •NO which, as already highlighted, in a spatially coordinated action with the high activity of Nox2 system, allows the reaction between •NO and O2•-, giving rise to ONOO-, a potent antimicrobial agent. Furthermore, •NO is an important signaling agent [41,42].
2.5.2. Phenotypic subpopulations of M2 MФs: cytoplasmic and extracellular components and reparative action (resolution of infection and healing)
2.5.2.1. MФs repertoire during tissue repair and regeneration/remodeling
Congruent with a population phenotype of M2 MФs, the highly elaborate machinery for the resolution of the infectious/inflammatory focus and tissue repair has as its main and distinct characteristic the production of high levels of Arg1 with the production of polyamines and specific amino acids. M2 MФs take part in tissue repair directly or by secreting paracrine factors, which will favor the healing process. The repair activity carried out by MФs is beyond the scope of this review and can be reviewed in detail in Das et al. [32]. Moreover, M2 MФs have the ability to suppress the proliferation of T cells associated with inflammation.
2.5.2.2. Role of extracellular matrix and adjacent cells
The role of the extracellular matrix has been highlighted during polarization. Components of the extracellular matrix, such as type 1 collagen, can modulate polarization [38]. In addition, studies have shown that differences between the cells present in each tissue/organ and the different cells recruited in different phases of the immune response can alter the polarization profile, through cellular communication with the production of substances that influence polarization programming [38]. As examples, the recruitment of fibroblasts during healing and the production of DAMPs, RAMPs, and SPMs can modulate polarization (Table 2) [38].
3. Immune response via polarized MФs against pathogens in general
In general, studies converge in pointing out that there is a transcriptional and metabolic/functional profile called the "common host response" to infection in which the M1 program is a majority over M2 in the acute phases of diseases and infections, partly justified by the microbicidal characteristics of the M1 MФs sub-population, tending toward balance or a greater number of M2 MФs in the phases of infection resolution and tissue repair [29]. For example, Mège et al. [65] showed that, in general, the majority polarization of M1 MФs is seen in gastrointestinal infections (for example, in Helicobacter pylori gastritis) and active Tuberculosis (TB), whereas the polarization of M2 MФs is better described in lepromatous leprosy (LLep), Whipple's disease and some localized infections (keratitis and rhinosinusitis). However, as yet to be discussed, this phenomenon is dependent on the specificity of the inducing stimulus (microorganism and other stimuli giving rise to the response, Table 2 and Table 3) and affected tissue, as well as the kinetic profile that is peculiar to each infection, whether acute or chronic. Thus, there are microorganisms that provide a specific context that contradicts this maxim, so that the M1 response may not be as intense and the M2 response may increase in the acute phase and therefore be protective, partly due to an M2 profile that includes signaling between innate and acquired immunity favoring the Th2 cell profile and/or due to the vital importance of the organ that is infected [29]. Therefore, the response pattern involving MФs commonly and formerly described as canonical may not occur as such either.
However, understanding this duality of inflammatory and anti-inflammatory responses in the context of lung infections seems to be increasingly necessary. For example, while certain infections are contained by a Th1/M1 MФs response and this response generates serious lung damage, sometimes may be more deleterious than the infectious disease itself (e.g. CoronavirusDisease-2019, COVID-19), Dolgachev et al. [66] showed that IL-10 overexpression in an experimental model of pulmonary contusion generates robust activation of M2 MФs and, in such a context, there is a lower intracellular microbicidal power associated with increased bacteremia, which accelerated mortality in pneumonia caused by Klebsiella pneumoniae. These observations regarding duality and the effective response without harming the host have an immediate impact, since they lead to the therapeutic use of anti-inflammatory drugs (such as corticoids) and antioxidants.
The context for understanding the polarization of MФs in infectious diseases in which pathogens show tropism for certain cells/tissues is more complex, as examples, pathogens that invade MФs, such as influenza and dengue viruses, intracellular bacteria such as Brucella spp., Salmonella spp, Listeria monocytogenes and Mycobacterium tuberculosis, fungi such as Candida albicans and Aspergillus fumigatus, and parasites such as Leishmania spp. They present, in this context, an advantage for modulating polarization and, consequently, the entire immune response following the first contact between the immune system and the microorganism, with the aim of replicating in these cells and consequently perpetuating the infection and/or establishing the disease [67].
Consequently, this review will also focus on the virulence mechanisms by which microorganisms can, at any time, alter the functional profile of MФs populations by modulating the production of local mediators and/or masking/altering the respective PAMPs, thus, for example, altering M1 to M2 polarization, or vice versa, and contributing with microbial survival and maintenance of the infectious focus. Table 4 presents a list of microorganisms that are potentially pathogenic to humans and which, according to the literature, can induce a well-defined polarization of MФs. These microorganisms are discussed further below, in terms of the peculiarities of inducing polarization and the mechanisms of virulence factors that can affect the protective response. The literature also points out that a given polarization of MФs can be beneficial, i.e. protective for the host, or deleterious, i.e. favor the outcome of the disease caused by the microorganism [29,65,68], so Table 4 also shows the majority protective profile.
3.1. MФs polarization in infectious diseases
The polarization of MФs can occur in the context of inflammatory-infectious and inflammatory-non-infectious diseases, the latter of which is beyond the scope of this review, so only the polarization of MФs induced by microorganisms will be addressed here. Therefore, the polarization of MФs against microorganisms that are potentially pathogenic to humans and relevant to clinical medicine is presented below, as summarized in Table 4.
3.1.1. Polarization of MФs in response to infectious diseases caused by bacteria
3.1.1.1. Gram positive bacteria
- Staphylococcus aureus
Since it was first reported in the literature in 1880 by Ogston [69], S. aureus has gained notoriety over the decades, mainly because of its versatility as a commensal of the human microbiome and, in certain predisposing conditions (such as immunodeficiencies, the presence of surgical wounds, certain comorbidities or infection by highly virulent or pathogenic strains of this bacterium), as a causative agent of various human pathologies [70]. Currently, about 20-80% of the human population has its microbiome colonized commensally by S. aureus [71]. Infectious diseases (skin, soft tissues, and invasive) caused by this microorganism represent a serious public health problem, because of healthcare settings and community-associated S. aureus potentially pathogenic [72].
The immune response to S. aureus varies considerably according to the anatomical site and time of infection, ranging from an M1/Th1/Th17 to an M2/Th2 cellular profile, and there are cases in which neutrophils play a sufficiently important role to contain the infection. In these contexts, HIF-1a is an important regulator of the immune response. However, the real protective role of MФs in infectious diseases caused by S. aureus remains undefined, since this microorganism can affect different organs and tissues and make use of a vast repertoire of virulence factors [73].
Recently, Xu et al. [74] reported that during pulmonary infection by S. aureus the protective response performed by alveolar MФs is M1, and the induction of Akt1 signaling reinforces the activity of SOCS1 and inhibits NF-κB, altering the functional microbicidal profile of the M1 MФs sub-population to a non-functional M2 phenotype. Later, using another model of deep tissue infection, Wang et al. [73] demonstrated that the response in liver infections by S. aureus seems to involve MФs-MOs and proinflammatory Kupffer cells (M1) associated with Th1/Th17 lymphocytes, which recognize PAMPs from the microorganism through the TRL2 receptor, this process being strongly regulated intrinsically by the FoxO1 transcription factor of the MФs. According to the authors, the high expression/activity of FoxO1 associated with TLR2 favors an M1 profile, whereas in mice deficient (knockout LysCre/+FoxO1fl/fl) for this transcription factor, the proinflammatory response is abolished, favoring an M2 sub-population profile, in the latter case, with an increase in the pathogenic potential of the bacteria. These findings demonstrate that an M1/Th1,17 type response is protective in invasive infections caused by S. aureus. Furthermore, Kasraie et al. [75] showed that the α-toxin produced by S. aureus induces a polarization related to the Th1 profile related to the production of the CXCL10 chemokine.
However, demonstrating the complexity of the protective response that can be developed against S. aureus, the microorganism can naturally produce a range of mediators (SEA, SEB, protein A, α toxin) that change the polarization profile of MФs [76,77]. In fact, these mediators can favor the progression of an acute infection to a chronic or associated with biofilms, so in the early stages of infection, a proinflammatory profile is observed, but this may not be effective in eliminating the microorganism, in addition to, if exacerbated, causing tissue damage in the host. Therefore, the subsequent recruitment of M2MФs also eliciting a Th2 response appears to be effective in eradicating the chronic infectious disease or biofilms [76,78]. Given this, Krysko et al. [79] showed that in chronic rhinosinusitis with nasal polyps caused by S. aureus there is a majority activation of M2 MФs, and these cells have little ability to phagocytize the microorganism and contribute to extensive microbial colonization. In addition, S. aureus is a known inducer of biofilms on material surfaces and in in vivo infections, and the intense activation of M2 MФs can occur with associated tissue fibrosis, due to an activation involving MyD88 signaling independent of TLRs [80].
- Listeria monocytogenes
Listeria monocytogenes is responsible for infections in humans with pathogenic potential to generate diseases such as meningitis. According to the literature, the response involving M1 MФs is protective against this microorganism, since it has been shown that mice experience persistent infection with extensive disease severity in the absence of adequate IFN-γ or TNF-α production [11,28,33,81].
- Streptococcus spp.
Streptococcus pyogenes and Streptococcus pneumoniae
The effective response against both S. pyogenes and S. pneumoniae involves the polarization of M1 MФs. In the clinical context of infectious diseases caused by these microorganisms, complex factors and modulators of polarization are present [29].
- Tropheryma whipplei
Tropheryma whipplei causes a rare systemic disease in humans, Whipple's disease [29]. Desnues et al. [82] demonstrated in vitro the microbicidal and protective profile of M1 MФs against T. whipplei. The profile of M2 MФs is associated with the chronic and systemic form of the disease, as well as the intestinal chronic form [83].
3.1.1.2. Gram negative bacteria
- Salmonella typhimurium
Infectious diseases and poisoning caused by species of the Salmonella genus remain a public health problem worldwide [29]. In addition, sepsis caused by S. typhimurium is another serious problem [84]. The protective response involves M1 MФs. However, the exacerbated response of this profile is seen in sepsis and, when uncontrolled, can increase morbidity and mortality [84].
- Escherichia coli
Escherichia coli is part of the human gut microbiome, as a commensal. In certain predisposing conditions and for the acquisition of certain pathogenic strains, varying degrees of illness can occur, from milder intoxications to more severe infections, where there is a high capacity for microbial multiplication and tissue invasion that can lead to sepsis. The different strains of E. coli can cause different MФs polarization and, as with other commensals, generate immunotolerance. Escherichia coli is one of the main microorganisms producing high levels of LPS, a PAMP that induces polarization of M1 MФs, as well as being one of the main molecules inducing sepsis [65]. Sepsis can occur during E. coli infection, i.e. a systemic inflammatory response syndrome (SIRS), involving M1 MФs [65]. However, when there is a favorable prognosis for the course of the infectious disease and, in certain cases, respecting the canonicity of the response involving these cells, the reprogramming of MФs to an M2 profile allows tolerance to LPS and cessation of sepsis, in what is known as LPS-reminiscent immune dysfunction [65]. Thus, strains of E. coli that produce high levels of LPS or that have a high proliferation rate (as the disease caused by this commensal can be quantitative), without prior immunotolerance, cause polarization of M1 MФs [65]. On the other hand, immunological tolerance to LPS can cause polarization of the M2 profile [65]. In patients with severe sepsis or the associated SIRS, pharmacological intervention with the clinical use of corticosteroids is necessary, aimed at decreasing the Th1/M1 response and increasing Th2/M2, otherwise in the absence of the compensatory anti-inflammatory response syndrome (CARS).
- Pseudomonas aeruginosa
Pseudomonas aeruginosa is the main etiological agent of lung infection in patients with cystic fibrosis (CF), with approximately 80% of CF patients having their lungs colonized/chronically infected with this microorganism, resulting in considerable morbidity and mortality rates [85]. During lung infection, the increase in mucous secretions in alveolar areas makes the microenvironment conducive to the formation of biofilms by P. aeruginosa [85]. Just as a more exacerbated M2 MФs profile is observed in biofilms caused by S. aureus [80], CF patients experiencing chronic infection by P. aeruginosa exhibit high levels of IL-5, IL-13 and IL-17, and an increase in eosinophils and an associated change in the polarization profile from M1 to M2 are also observed, being influenced by the formation of biofilms [85]. Furthermore, in the protective response against localized keratitis caused by P. aeruginosa, there is an increase in IL-33, a cytokine member of the IL-1 family, which amplifies the response involving M2 MФs [65]. The genetic background of the individual is essential in the defense against this disease, since it has been reported that BALB/c mice, which tend to have a more pronounced Th2/M2 response than Th1/M1, are resistant to keratitis caused by P. aeruginosa, while C57BL/6 mice are sensitive to the disease [86].
- Chlamydia spp.
Species of the genus Chlamydia can cause lesions in the eye and genitalia and can affect anatomical structures that compromise fertility and women's health. It is considered an sexually transmitted infections (STI) of importance when it comes to implementing public health actions [87]. Chlamydia trachomatis is an obligate intracellular bacterium that infects the ocular epithelium and the genitourinary system, causing trachoma, a considerable cause of preventable blindness, and damage to the reproductive system, respectively [87]. The polarization of M2 MФs, associated with a Th2 profile (increase in IL-24), in response to C. trachomatis infection, has been speculated as an appropriate coevolutionary response between pathogen and host in a context that involves protection against the pathogen and concomitant immunopathological non-aggression against structures vital for human reproduction, logically in the light of evolution that by selection favors reproduction and, since a majority and persistent response involving M1 MФs can damage such structures, M1 MФs would only occur to a lesser extent although still effective in protection [33,87,88]. This may be explained by the large number of asymptomatic infected individuals [87]. Furthermore, ex vivo studies show that T cells stimulated with antigen from C. trachomatis secrete IL-4, and not IL-17 or IFN-γ, confirming the Th2 profile [87].
As for Chlamydia pneumoniae, which causes Chlamydia pneumonia and is responsible for exacerbating asthma and chronic obstructive pulmonary disease (COPD), Jupelli et al. [89] showed that although M1 MФs are effective in eliminating the pathogen, the response is exacerbated, causing excessive inflammation, lung injury and fibrosis (related to poor signaling via IL-1). This profile of an exacerbated response involving M1 MФs against pulmonary pathogens has been firmly established in recent literature, and such a response can lead to greater tissue damage and increase the death rate associated with the disease, as is being evidenced in COVID-19 patients [90].
- Brucella spp.
Brucellosis caused by Brucella spp. is a widespread zoonotic disease worldwide [91,92]. The response involving M1 MФs is protective for the host, while the bias of the response toward a predominantly M2 pattern, via an increase in the microorganism's Omp25, or the activation of PPARγ by other sources, favors brucellosis [29,91].
- Shigella flexneri
Shigella flexneri can induce apoptosis in MФs activated by INF-γ [93]. In addition, the response via M1 MФs is protective, and virulence factors of the microorganism causing a bias toward the M2 profile favor the disease [94].
- Coxiella burnetii
Coxiella burnetii is an obligate intracellular bacterium that causes an acute disease known as Q fever, with a risk of chronic evolution of fever in immunocompromised patients, based on the interaction with MФs [29]. The lack of canonicality in the dynamics of the M1 and M2 MФs profiles can occur in patients with Q fever in the early stages of the disease and later, requiring a peculiar diagnosis and therapy aimed at a more favorable prognosis, in which the persistence of the M2 profile has been reported to be associated with the continuity and severity of the disease [95].
- Yersinia enterocolitica
Species of the genus Yersinia, including Y. enterocolitica, are facultative intracellular pathogens for humans that can induce apoptosis in circulating MФs [93]. The response involving M1 MФs has been reported to be protective, while the microorganism-induced switch to the M1 program favors disease [29].
- Francisella spp.
Tularemia is a febrile disease caused by bacteria of the genus Francisella, mainly F. tularensis. The form of pneumonia induced by this bacterium has a delay in the innate immune response of around 48-72 hours, followed by an influx of phagocytes and a cytokine storm [96]. Defects in M1 activation and predisposition to exacerbation of the M2 profile have been reported as risk factors for the severity of the disease [28,96].
- Helicobacter pylori
Helicobacter pylori infects half of the human population and can, in certain cases, cause peptic ulcers, chronic gastritis, and chronic infection-associated cancer (adenocarcinoma) [30,97]. During infection by H. pylori, in addition to the regulation of the response involving a mixture of the Th1/Th17 response, the participation of M1 MФs is evident, with concomitant participation of regulatory MФs (Mreg), and also association of M2 MФs (involving HO-1), showing the complex mix of MФs and the non-canonical context of polarization involved in infection by this microorganism [97]. Mutant mice with HMOX-1 deletion increase gastritis associated with Th1/Th17/M1 response [97]. In fact, studying the still undefined role of polarization in H. pylori infections, previous findings by Quiding-Järbrink et al. [30] already indicated the participation of the M1 MФs response, polarization that can be accelerated by prior vaccination. The timeless mixture of M1 MФs and M2 during uncomplicated H. pylori infection was also highlighted by the authors, with emphasis on the exacerbation of the M1 profile when severe gastritis occurs (with complications), a precancerous lesion.
- Vibrio cholerae
Khan et al. [98] showed that V. cholerae, the causative agent of cholera, through its porin OmpU, induces a strong polarization of M1 MФs via interaction with TLR1/TLR2, recruiting MyD88 which culminates in the activation and translocation of the p65 subunit of NF-қB, despite the immunomodulatory duality of OmpU as a PAMP, also activating M2 MФs.
- Haemophilus spp.
Bacteria of the Haemophilus genus, causative agents of STIs (Haemophilus ducreyi) and community-acquired pneumonia (Haemophilus influenzae), are also responsible for intermittent infections that are one of the main causes of acute exacerbations of COPD [68,99]. A balance in the response of M1 and M2 MФs has been proposed in the course of infection by bacteria of this genus, both profiles being essential for controlling the disease [68,99].
- Borrelia burgdorferi
Borrelia burgdorferi is the causative agent of Lyme disease, with inflammatory arthritis (with a predominance of neutrophils) and heart disease (with a predominance of MФs) being the main outcomes of this disease [100]. The polarization of MФs should be interpreted with caution, since in vitro data show that the microorganism and its lipoproteins induce a clear M1 polarization with the production of proinflammatory cytokines (IL-1, -6, -12, and TNF-α) and •NO [100,101,102]. However, predominance or balance involving M2 MФs may occur in vivo [100]. This can partly be explained by the duality of the immune system in having to attack the microorganism in the context of Lyme disease while at the same time having to preserve important anatomical sites such as the joints [100,103] and vital ones such as the heart [100,104], so there is a balance or predominance of M2 MФs in these microenvironments.
- Neisseria gonorrhoeae
Neisseria gonorrhoeae, a facultative intracellular bacterium, is the etiological agent of gonorrhea. It is a considerable microorganism that causes STI, with approximately 100 million new infections every year worldwide, affecting groups that practice risky behaviors and occurring as a comorbidity [105]. Some factors make N. gonorrhoeae infection a serious public health problem, such as the lack of vaccines, resistance to antimicrobials and the absence of an effective human immune response. In fact, infection with this microorganism is incapable of inducing a protective response in the host, so that infected individuals develop gonorrhea, and it can occur repeatedly [105]. In part, this is due to evasion mechanisms developed by the microorganism, evading an innate and acquired immune response, despite a possible Th17 polarization and suppression of the Th1/Th2 response, and a varied polarization profile of MФs with an associated broad cytokine profile, which can be modulated by the microorganism to an M2 profile, which is not protective during infection [105].
3.1.1.3. Potentially pathogenic microorganisms in the oral cavity (Gram positive or negative)
Bearing in mind that potentially pathogenic microorganisms in the oral cavity are normally the commensals of this microbiome, which benefit from predisposing conditions in the host, these conditions being risk factors for the development of the respective disease, the coevolutionary context of the microorganism-host interaction tends to promote immunomodulation that can change in the context of the disease. In fact, different potentially pathogenic microorganisms belonging to the oral microbiome tend to promote, in vitro, a response involving M2 MФs, which are non-microbial and anti-inflammatory [21].
- Aggregatibacter actinomycetemcomitans
Aggregatibacter actinomycetemcomitans is a commensal of the oral microbiome and a potential periodontopathogen, representing one of the microorganisms involved, to varying degrees, in periodontal disease [21]. The immune response in the context of the disease, from acute episodes to chronic periodontal disease, involves a complex context in the pathogen-host interaction, with the immune response being complex and, in certain cases, related to levels of pathogenicity [21]. In vitro data reveal that the response involving the polarization of MФs seems to be related to the M1 profile, in direct pathogen-cell interaction, without considering the complex microenvironment of the oral cavity and deep periodontal tissues [21].
- Porphyromonas gingivalis
Porphyromonas gingivalis is a commensal of the oral cavity and potential periodontopathogen, which generates a polarization of M1 MФs, in a direct interaction involving microorganism-MOs in vitro, without considering the in vivo microenvironment [21]. Considering the in vivo microenvironment, interestingly, Papadopoulos et al. [106] showed that the challenge of precursor cells (MOs) with P. gingivalis made M2 MФs respond more intensely to the microorganism and produce high levels of TNF-α, IL-6, MCP-1 and MIP-1, compared to M1 MФs, including up-regulating the activation of T cells. On the other hand, M1 MФs produced higher levels of IL-10 than M2 MФs. Supporting these findings, the authors showed that the response of MФs M0 and M2 is associated with greater expression of TLR2, which is associated with a proinflammatory response. Furthermore, M2 MФs bound more robustly to P. gingivalis, being more effective in the response to the microorganism, in a context of the transition of the immune response and the cytokine profile involving the Th response common in periodontal disease.
- Fusobacterium nucleatum
Representing another commensal of the oral cavity microbiome, F. nucleatum generates a mixed polarization of M1 MФs and M2 in vitro during direct microorganism-MOs interaction [21].
- Prevotella intermedia
A mixed polarization involving M1 MФs and M2 is also observed in the direct interaction between Prevotella intermedia and cells, in vitro [21].
- Streptococcus spp.
Streptococcus spp., including Streptococcus mutans, Streptococcus gordonii, and Streptococcus sanguis, are microorganisms belonging to the microbiome of the oral cavity, which can, under predisposing conditions, participate in and trigger pathological processes, causing diseases, such as, periodontal disease and caries. Huang et al. [21] reported that these microorganisms can generate primary polarization, in vitro, involving M2 MФs.
3.1.1.4. Mycobacteria
- Mycobacterium tuberculosis
Mycobacterium tuberculosis (Mtb) is the main species belonging to the Mycobacterium tuberculosis complex and is the main etiologic agent of pulmonary tuberculosis (TB), considered one of the biggest causes of death from infectious diseases worldwide. Tuberculosis is still an infectious and contagious disease that has been declared a serious public health problem worldwide, especially in underdeveloped countries, with an estimated quarter of the world's population infected with Mtb and at risk of developing the disease, with approximately eight million new cases and almost three million deaths annually. In Brazil, TB has a considerable prevalence and is responsible for a significant rate of mortality and DALYs/YLDs, especially in people living with HIV-1 (PLHIV-1) during co-infection and in patients with comorbidities that affect the immune system [20,107,108,109].
In this context, among the 1/4 of the world's population infected with Mtb, around 10% will develop the disease at some point in their lives, due to possible immunosuppression (such as an altered M1/N1/Th1→M2/N2/Th2 pattern) [107]. After infection with Mtb, the target cells of the mycobacteria are the alveolar MФs, which in turn recognize the mycobacteria, promoting the immunomodulation of polarization and consequently of the associated Th profile. Thus, the control of polarization is an important target for the success of the mycobacterium, since this phenomenon controls the antimycobacterial response, the formation of granulomas and the transition between innate and acquired immunity, thus greatly influencing the patient's clinical outcome [20]. Several microorganisms, including facultative intracellular mycobacteria, induce a transcriptional and functional profile of the common host response with activities belonging to the M1 program, with the polarization of classically activated MФs (M1, proinflammatory and microbicidal) in a host protective profile [14], encompassing a context of a balanced Th1 response activated by IL-12 and an increased stimulation by interferon (↑IFN-γ) [107,110,111]. Roy et al. [111] showed that the transcriptional factor Batf2 is involved in the activation of M1 MФs, as well as for PAMP (LPS), being responsible for the intense inflammation associated with the M1 profile in active infection in vivo. In contrast, susceptibility to Mtb seems to involve a range of signaling induced by the mycobacteria, favoring M2/Th2 polarization (increased levels of IL-4 and IL-13) [20,107], with high levels of IL-10 being seen in active TB [112]. Furthermore, unlike in vivo microenvironments, in an in vitro model the stimulus with Mtb per se appears to activate M2 MФs, through the overexpression and activation of PPARγ linked to the mannose receptor, a situation that can occur in vivo, in the context of active TB, when mycobacteria are favored by virulence mechanisms [113], as will be discussed later.
Thus, Mtb and other mycobacteria that can also cause disease in humans generally elicit a protective response via M1 MФs [107]. Kahnert et al. [107] showed the protective action of M1 MФs against Mtb, while a change to a profile of M2 MФs favors active TB. Furthermore, vaccination with Bacille Calmette-Guérin (BCG) increases the activation of M1 MФs and guarantees greater protection in terms of preventing active TB [65].
Evidencing the importance of the Th1-type immune response and polarization of M1 MФs against Mtb and, since Mtb/other microorganisms co-infection is a very common event in the clinic of infected patients, Aira et al. [114] showed that a previous exposure of APCs and MФs to antigens from Hymenolepis diminuta and Trichuris muris led to an anti-inflammatory type response at certain stages, which altered the polarization profile toward M2 MФs, thus decreasing the response to mycobacteria during co-stimulation. On the other hand, Mtb co-stimulation and prior exposure to Schistosoma mansoni promoted a synergistic response involving the M1 MФs program, which favored the elimination of mycobacteria through phagocytosis and intracellular death by mycobactericidal mechanisms of the phagocytes.
As has been observed for chronic infectious diseases in general, including TB, a natural variation between the sub-populations of MФs in active TB may be related to the kinetics and specificity of a constant host response to the pathogen (proinflammatory action) and the associated need to repair lung damage (anti-inflammatory action) [29]. During granuloma formation in active TB, certain levels of ATP (100 µM) can lead MФs to a momentary polarization toward an M2-like anti-inflammatory state, due to ATP hydrolysis mediated by CD39 generating AMP, with the involvement of A2A receptors [84]. In addition, Mtb and other microorganisms (Leishmania spp., Legionella pneumophila, Trypanosoma cruzi and Toxoplasma gondii) can generate AMP through ATPase activity, also causing a momentary change in polarization toward M2 MФs [84].
- Mycobacterium leprae
Mycobacterium leprae, an intracellular pathogen that preferentially infects MФs (DCs-MOs), is the etiologic agent of leprosy, a debilitating chronic infectious disease with a high lethality rate [115,116]. In this disease, a diverse repertoire of immune responses stands out. In tuberculoid leprosy (TLep) there is a predominance of M1 MФs, in which high levels of IFN-γ and IL-5 prime MФs for this profile, while in LLep a predominant profile of M2 MФs (ILs 4 and 10) associated with humoral immunity is observed [115]. Evidencing that the response involving M1 MФs is protective, it is highlighted in the literature that prior vaccination with BCG can preferentially direct an effective and sustained M1-type response, which protects the host against M. leprae and prevents the development of the disease [115,116]. In addition, Th1 cytokines are known to induce macroautophagy, a mechanism that has been shown to protect against leprosy [116]. Mycobacterium leprae/HIV-1 co-infection can alter the expression of cytokines and chemokines and the polarization profile of MФs during the interaction [117].
3.1.2. MФs polarization in infectious diseases caused by fungi
- Candida albicans
Candida albicans is a commensal yeast of the human microbiome, which can cause opportunistic infectious diseases when there are favorable conditions for the microorganism, such as immunosuppression, the presence of unhealed wounds and a decrease in the number of bacteria in a given microbiome [118,119,120,121]. Some C. albicans PAMPs recognized by PRRs activate NF-kB and ↑ proinflammatory cytokines, favoring the M1/Th1 type host protective response, as shown in Table 4. On the other hand, representing a new role for MФs in gastrointestinal and systemic diseases caused by C. albicans, according to Lefèvre et al [122], expression of dectin-1 and other mannose receptors in MФ can, in certain contexts and in a scenario involving IL-13, generate the induction of alternatively activated MФs (M2), which can also favor fungicidal activity aimed at resolving the infectious focus. However, in a study involving a complete proteomic characterization of the MФs response to a strain of C. albicans isolated in the clinic (SC5314), Reales-Calderón et al. [123] reported the importance of M1 MФs in effectively fighting the microorganism, in contrast to the M2 MФs profile, which seems to favor the survival and dissemination of the fungus and lead to greater pathogenicity associated with the infection. The authors showed that different structural and functional components (involving metabolic and immunological programs and their mediators) are differentiated in the M1 and M2 subpopulations of MФs.
- Pneumocystis jirovecii
Pneumocystis jirovecii is an opportunistic pathogen, a cause of Pneumocystis pneumonia with a serious risk of death in immunocompromised patients. As with the most varied infectious diseases of the lung, although the microbicidal/inflammatory process is essential to fight the pathogen, the excessive response can damage the organ, thus also causing high morbidity and mortality in the context of the infection [124]. Alveolar MФs recognize PAMPs in P. jirovecii and participate in the primary response to infection, and it has been shown that these cells can stimulate a balance between a pro- and anti-inflammatory response, with activation of both profiles, M1 and M2, with production of IFN-γ, but the response via activation of the M2 profile generates higher levels of IL-4 generated by CD4+ T-cells and a more effective response, which neutralizes and promotes clearance of the microorganism more adequately and without damaging the lung [124]. The authors also point out that antibiotics can amplify the proinflammatory damage and that corticosteroids can attenuate this damage and contribute greatly to the survival of critically ill patients. The authors also highlighted the plasticity of the response involving MФs.
- Cryptococcus neoformans
The most effective immune response to C. neoformans, the causative agent of cryptococcosis, seems to involve Th1 signaling associated with the classic activation of MФs (M1) with considerable fungicidal capacity [125]. This profile shows a marked presence of T effector cells (CD4+/Th1 and CD8+) and M1 MФs, high levels of IFN-γ, TNF-α and IL-12, marked production of •NO, culminating in complete clearance of the microorganism in the lung, as opposed to the Th2 response, involving B cells (Th2/humoral response with high levels of IL-4, -5, -10 and -13) and M2 MФs, with marked induction of arginase, which consumes arginine without producing •NO, facilitating microbial dissemination/CNS infection and increasing lethality [125,126]. Eastman et al [126] showed that virulence factors in C. neoformans, such as SSA1 expression and associated production of Ssa1 (a HSP70 homologue), favor the initial transition of the M1 MФs response to a majority M2 MФs profile, even during primary lung infection. Although M2 MФs preserve lung tissue, this transition (M1→M2 MФs) favors dissemination and lethality in the early stages of infection. For example, it contributes to cerebral cryptococcosis. This early virulence factor is added to others already identified at other stages, such as laccase production, increasing fungal survival and favoring disease outcome.
- Aspergillus fumigatus
Aspergillus fumigatus and some other species of the Aspergillus genus are potentially pathogenic fungi, mainly affecting patients with immunodeficiencies related to the quantity and/or dysfunction of phagocytes, predisposing the infected to pulmonary and systemic pathologies [2]. It is widely known that patients with a quantitative decrease in phagocytes are more predisposed to developing systemic aspergillosis, while those with some qualitative dysfunction of phagocytes (such as CGD or G6PD deficiency) are more predisposed to chronic local infections aggravated by granulomas [127]. In the context of MФs polarization, the M2 profile seems to protect the host without causing detriment to the pulmonary microenvironment [28].
- Paracoccidioides brasiliensis
Paracoccidioides brasiliensis (Pb) causes paracoccidioidomycosis (PCM), a disease that has a considerable incidence in Brazil. Recently, Freitas et al. [128] demonstrated that paracoccin, a Pb PAMP, can be recognized by TLR4 receptors on MФs and generate M1-type polarization, which has a strong protective effect against PCM. Confirming the functional profile of MФs against Pb, da Silva et al. [129] showed that extracellular vesicles (EV) secreted by the fungus also induce M1-type polarization. The authors suggest that these findings may provide an opportunity to use PAMP or EV components to boost the immune response of MФs against Pb, in the search for new therapeutic options to treat PCM.
3.1.3. MФs polarization in infectious diseases caused by parasites
- Plasmodium spp.
Malaria is still a neglected disease, despite its magnitude, especially in underdeveloped areas and countries [84]. In Brazil, there are endemic areas that represent a serious public health problem, with Plasmodium falciparum being one of the main etiological agents of the disease. During active disease, the MФs profile seems to predominate, while a balance of response (M1 = M2 MФs) must be achieved with a view to resolution in immune/asymptomatic individuals and in the treatment of malaria [130]. As for Plasmodium berghei, the high degree of plasticity of MФs reprogramming confers a unique context during infection and disease, for example in the lung, ensuring a marked MФs response during active disease [130].
- Toxoplasma gondii
Toxoplasma gondii is an obligate intracellular parasite, propagating in blood cells and reaching vital organs such as the CNS, being the causative agent of toxoplasmosis [131]. The main immune response to the parasite has been reported as belonging to the Th1/M1 program, and has had repercussions in different studies on the gestational aspects of the host's response to the parasite, since the infection of pregnant women by T. gondii and the associated deleterious effects on the embryo/fetus is considered a serious public health problem, due to vertical transmission through the placenta [52,131,132].
- Trypanosoma cruzi
Chagas disease, whose etiologic agent is T. cruzi, is still considered a serious public health problem in Brazil and most Latin American countries, affecting around 7 million people worldwide. Cardiovascular complications, including Chagas cardiomyopathy, which affects around 30% of those chronically affected by the disease, are the most serious repercussions and cause considerable sequelae and DALYs/YLDs [133,134]. In the context of the response involving phagocytes, M1 MФs predominate during the acute phase and M2 during the chronic phase. The predominance of M1 MФs with a high microbicidal capacity, if effective, can favor the host, since the parasite load is considerably reduced in the myocardium in the early stages, thus preventing the disease from becoming chronic [133,135]. However, modifications to the response that generate a change in polarization in the early stages of the disease can determine the severity and chronicity of the infection [133]. Ponce et al. [133] showed that CD73, an enzyme that cleaves ATP and generates extracellular adenosine, can attenuate inflammation and afford the deactivation of M1 MФs. In this sense, CD73 inhibition decreased IL-4 and IL-10 levels and restored the M1 profile, resulting in a decrease in serum brain-muscle creatine kinase isoenzyme and improvements in electrocardiographic characteristics during the chronic phase.
- Leishmania spp.
Infectious diseases caused by species of the genus Leishmania are endemic in some regions, including some regions in Brazil. After infection, M1-programmed DCs-MOs in skin tissues are the main effectors of the immune response in humans against the parasite, leading to a Th1-type response that protects the host. Species of the Leishmania genus are obligate intracellular parasites of neutrophils or, preferably, MФs [136]. Once phagocytized, the parasites (Leishmania major, Leishmania donovani, and Leishmania braziliensis) are eliminated intracellularly through the expression of microbicidal factors present in M1 MФs, such as the expression of iNOS and the production of •NO. On the other hand, the development of Leishmania spp. in M2 MФs favors the parasite and its cycle, since, in these cells, the expression of Arg1 and action on arginine, generating ornithine and, subsequently, promoting the synthesis of collagen, as well as polyamines, favors the growth and proliferation of the parasite [136,137]. At the same time, neutrophils represent the first line of defense in infections by species of the Leishmania genus. Guimarães-Costa et al. [59] showed that NETs produced by neutrophils can repress the polarization of the M1 MФs subpopulation, aggravating leishmaniasis.
- Cryptosporidium parvum
Cryptosporidiosis in Brazil has a considerable prevalence, especially that caused by C. parvum. The effective response involves polarization of M1 MФs [11].
- Schistosoma spp.
Schistosomiasis, caused by species of the genus Schistosoma, is a zoonotic parasitic disease that is considered to be the second most prevalent worldwide, with around 200 million people infected, and is a serious public health problem in precarious areas [84,138,139,140,141]. Schistosoma mansoni is responsible for a considerable prevalence of schistosomiasis in Brazil, especially in areas with poor basic sanitation and which are used for recreation/activities with standing water, where snails of the Biomphalaria genus are found [140]. The M1-type response may be the majority in active schistosomiasis in early stages of intense inflammation [114], in a context where this exacerbated response promotes deleterious tissue damage in the liver and other organs, such as the intestine and lung, and considerable development of sequelae that lead to physical limitations in patients affected, impacting on DALYs/YLDs [138,139,141]. However, studies indicate that the response presents a profile in the initial stages with a predominance of M1 MФs (increase in IL-12 and expression of iNOS), changing to polarization in M2 MФs (increase in IL-4 and IL-10 and expression of ARG-1) after the production of parasite eggs, due to the change in antigens presented [138,141]. In this context, in which the absence of IL-4 signaling can increase lethality in schistosomiasis, after egg-laying, this interleukin promotes Th2/M2 polarization in order to promote tissue repair and limit tissue injury, but leads to liver fibrosis [138,141]. Therefore, the balance between the Th1/M1 and Th2/M2 responses is fundamental for protecting the host.
- Other helminths
Infection by helminths and associated diseases affects a considerable portion of the world's population, especially in underdeveloped areas with poor sanitation, with considerable morbidity and mortality rates, as well as having an impact due to DALYs/YLDs [84]. As with the other parasites mentioned so far, a majority response involving a certain population phenotype of MФs depends on several factors, such as: the stage of the parasite's life cycle; the presence or absence of virulence factors as modulators of the response; the affected organ; and host control. Even so, the canonical pattern may not occur, so there are still several gaps in the understanding of polarization in the context of parasite infections. Among the parasites that trigger a majority polarization of the Th2/M2 MФs type, regardless of the occurrence or not of canonicity, with release of IL-4, IL-13, IgE and intense eosinophilia, can be mentioned: Ascaris lumbricoides, Trichuris trichiura, T. muris, Taenia crassiceps, Necator americana, Ancylostoma duodenale, Nippostrongylus brasiliensis, Mesocestoides corti and H. diminuta [130,142,143]. This tendency to polarize toward the Th2/M2 profile, even in an IL-4/IL-13-free microenvironment, with the involvement of B cells producing antibody, has in part been described as being dependent on the PAMPs present on the surface of the parasites, such as glycans, including Lewis X, LDN, LDN-DF, fucosylated structures such as chitin, tyvelose and trehalose, which are recognized by CLRs and TLRs [144]. Evidencing this fact, Tawill et al. [145] showed that filaria extracts with enzymatic removal of glycans led to an absence of activation of the Th2 profile response.
3.1.4. Polarization of MФs in response to infectious diseases caused by viroses
- HIV-1
Since the beginning of the 1980s, when the pandemic/epidemic began, the spread of HIV-1 infection and the increase in the number of PLHIV-1/AIDS continue to be a serious public health problem in Brazil [146,147]. This retrovirus has a single-stranded RNA genome with positive polarity [(+)ssRNA] and belongs to the Retroviridae family [148]. HIV has an avid tropism for CD4+ T lymphocytes, the population of lymphocytes that takes the greatest part in polarization, and the virus presents important PAMPs that guide the polarization of MФs, including the gp120, Tat, and Nef proteins that induce the activation of M1 MФs [149]. However, in the context of the immune response developed by MФs, Cassol et al. [150] demonstrated that both sub-population profiles, M1 and M2, are important for the host response to HIV-1, but that the prevalence of a sub-population has an impact on specific stages of the viral replication cycle. Thus, in general, a balance between the M1 and M2 response occurs (Table 4), with changes in profiles in temporal functions of the orchestrated response to the pathogen. In vitro, HIV-1 directs a polarization of MФs with M1 programming, with marked production of cytokines and chemokines of type 1 inflammation, increased oxidative burst and •NO production, and this program seems to be the most potent antiviral [84]. Thus, a persistent and exacerbated population profile of M2 MФs may correlate with the severity of the disease, in which the HIV-1 cycle seems to be slightly favored by the population increase of MФs of the M2 program [20,33]. Furthermore, the control by HIV-1 of the alternation of subpopulations during infection may be related to greater cellular permissiveness to the fullness of the viral cycle and the dissemination and establishment of AIDS [148].
Galvão-Lima et al [149] reported that MФs from PLHIV-1on highly active antiretroviral therapy (HAART) compared to those from PLHIV-1 not on HAART differ in their functional profile and ability to react to different stimuli, including LPS.
- HTLV-1
The human T-cell lymphotropic virus (HTLV) includes two main subtypes, HTLV-1 and HTLV-2. HTLV-1 predominantly infects CD4+ T lymphocytes [5]. A considerable portion of the world's human population is infected with this virus asymptomatically, however, chronic manifestation of the disease can occur, as well as co-infection in PLHIV-1 by intracellular pathogens, with HTLV-1/Mtb and HTLV-1/HIV-1 co-infections being common [5,151]. It should be considered that the imbalance in an optimal immune response to various pathogens, including the activation of MФs polarization, caused by the interference of HTLV-1 infection and its tropism for CD4+ T lymphocytes, has been extensively reported [152,153,154,155,156]. In a study carried out by Amorim et al. [5], it was observed that MФs isolated from people living with HTLV-1, when compared to control groups (including uninfected individuals), present a similar ability to kill L. braziliensis, however, higher levels of CXCL9 and CCL5, and lower levels of IL-10 were detected. There were no differences in IFN-γ levels, which partly explains the similar ability of the groups to control the parasite, since the effective intracellular response against L. braziliensis mainly involves M1 MФs activated by this cytokine. To explain these findings, a classic event has been reported, in which the pathogenesis of the disease involving infection of CD4+ T cells by HTLV-1 is characterized by uncontrolled proliferation of these cells and consequent activation of the Th1 profile with associated high production of TNF, IFN-γ, CXCL9 and CXCL10, as well as spontaneous activation of neutrophils with the development of an intense oxidative burst [5].
- HCV
Hepatitis C virus (HCV) infection can result in persistent liver disease in the majority of those infected, partly due to the virus's varied strategies for evading the immune system, which can lead to chronic hepatitis, cirrhosis and even hepatocellular carcinoma [157]. In the successful containment of HCV infection, seen in the minority of cases (up to 20% of those infected in the acute phase), the initial profile involving the activation of Kupffer cells and the recruitment of circulating MOs, which play both roles by differentiating into M1 MФs with an increase in IFN-γ by the Th1 profile, is essential for the proinflammatory defense against the virus and, followed chronologically by a change to an M2 MФs profile for complete viral clearance, caused by IL-4 and IL-13 from the Th2 program with production of the anti-inflammatory mediator IL-10, which promotes the resolutive phase with tissue repair [157]. However, as will be discussed later, the virus manipulates polarization with a view to the persistence of the disease, causing successive proinflammatory foci followed by repair, which generates continuous replacement of the original liver tissue with fibrotic tissue, as seen in liver scars [157].
- H5N1
H5N1 was responsible for a pandemic in 2009. The host's protective response to H5N1 involves a polarization of M2 MФs, with ↑ M1 MФs favoring the increase in viral load and disease progression [33,158,159].
- SARS-CoV and SARS-CoV-2
Severe acute respiratory syndrome coronavirus (SARS-CoV) is a virus with a non-segmented single stranded RNA genome and positive polarity [(+) ssRNA], belonging to the Coronaviridae family. SARS-CoV-2 is responsible for the recent pandemic, which claimed the lives of millions of people, promoting, in addition to asymptomatic or presymptomatic infection, varying degrees of disease intensity (COVID-19, mild illness, moderate illness, severe illness, and critical illness) [1]. The cytokine storm, the action of phagocytes during innate immunity and the profile of the acquired immune response are variable and impact disease outcomes, and the participation of MФs in COVID-19 raises the possibility of a plausible pharmacological target [33,90,163]. The great variation in the immune response to SARS-CoV-2 and the different disease outcomes is justified by the individual variations of the human immune system [164].
The initial infection of epithelial cells in the respiratory tract and oral cavity leads to the recruitment of MOs and MФs, which are subsequently activated. Prior to the current pandemic, the participation of MФs, especially alveolar MФs, in defense and clearance had already been highlighted in the response to SARS-CoV [148,165,166,167]. Before the COVID-19 pandemic, it was shown that at a certain point in the infection, SARS-CoV can induce intense polarization of M1 MФs through a cytokine storm and proinflammatory factors and a high influx of lymphocytes and dependence on antibodies, causing, due to this intense response, a notorious death of the MФs population, in this context forcing immunosuppression and/or immunopathology, therefore facilitating viral dissemination, the persistence of the infection and leading to a high lethality rate associated with the disease [148,165,166,167]. However, until now, the impacts of an exacerbated M1 MФs response reflecting in damage that leads to more severe clinical outcomes had been neglected.
As previously raised in SARS-CoV infections, the participation of MФs during SARS-CoV-2 infection remains controversial. In fact, this participation is factually contextual, depending on numerous host factors (affected tissue, MHC immune profile, immunological predisposition, etc...) and SARS-CoV-2 virulence factors [164], in line with the complex pattern of effective host defense against the virus seen in clinical setting. Thus, for example, since the beginning of the COVID-19 pandemic it has been speculated that, during the proinflammatory phase due to the intense microbicidal response of phagocytes performing type 1 inflammation (i.e., with increased oxidative burst, NET release, cytokine storm that amplify inflammation and favor clots), it would be ideal to modulate the M1 to M2 polarization as well as attenuate the inflammation that favors tissue damage and unfavorable outcome in COVID-19, for example, the clinical use of antioxidants has been speculated [90]. Lian et al. [161] showed in vitro that both M1 and M2 MФs are effective in fighting SARS-CoV-2, but only M1MФs lead to exacerbation of lung cell damage due to the intense inflammatory process. The Table 4 complements the current findings on the polarization of MФs in infections caused by SARS-CoV and SARS-CoV-2.
- RSV
Respiratory syncytial virus (RSV) is a single-stranded RNA [(-)ssRNA] genome virus belonging to the Paramyxoviridae family [148]. A summary of the polarization of MФs in the response to RSV is presented in Table 4, being M1-like protective in certain contexts.
- Coxsackievirus, group B type 3 (CVB3)
Coxsackie virus, or Coxsackievirus, belongs to the enterovirus family, has a broad tissue tropism and is a common cause of infection in newborns [168]. A summary of the polarization of MФs in infections caused by CVB3 is presented in Table 4, being M2 MФs more effective.
- HCMV
The human cytomegalovirus (HCMV) is a diverse β-herpesvirus that infects between 50-90% of adults, with tropism for various cell types, which can culminate in systemic conditions with a wide range of affected organs. The infection is mostly asymptomatic, although moderate to severe symptoms may occur, as in mononucleosis, and it is also associated with chronic inflammation such as arthrosclerosis [169]. The control of infection and disease caused by HCMV follows the classic model of MФs action, with a proinflammatory M1 MФs profile during the acute phase, followed by a reprogramming/transition to an anti-inflammatory M2 MФs profile during resolution and tissue repair. However, as a virulence mechanism mediated by NF-κB and activity of a specific kinase, when infecting a MФ, HCMV can alter phagocyte reprogramming and modulate M1 MФs ↔ M2 MФs profiles, aiming to escape host immune mechanisms as well as dissemination of the infectious focus [169].
- DENV
Dengue virus (DENV) is a single-stranded RNA genome virus with positive polarity [(+)ssRNA] belonging to the Flaviviridae family [148]. A classic quasi-equilibrium response involving MФ M1 and M2 is observed. Healthy individuals infected with DENV may show a balance involving a slight increase in MФ M1 in the acute phase, followed by an intense increase in the response involving MФ M2 [170]. However, Lee et al. [170] showed that a decrease in the response involving MФ M1 and M2, subsets M2a and M2b, tends to develop the hemorrhagic form of the disease, since these subsets of MФ M2 are related to type 2 inflammation and resolution of the infection/immunity transition.
Table 4.
Polarization of macrophages (MФs) against potentially pathogenic microorganisms of medical relevance.
Table 4.
Polarization of macrophages (MФs) against potentially pathogenic microorganisms of medical relevance.
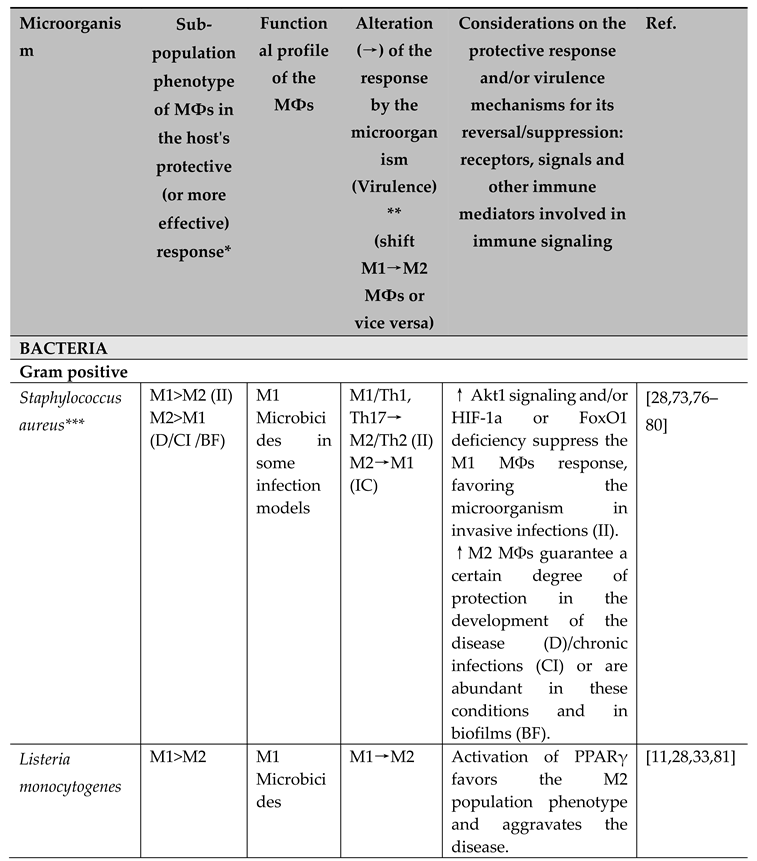 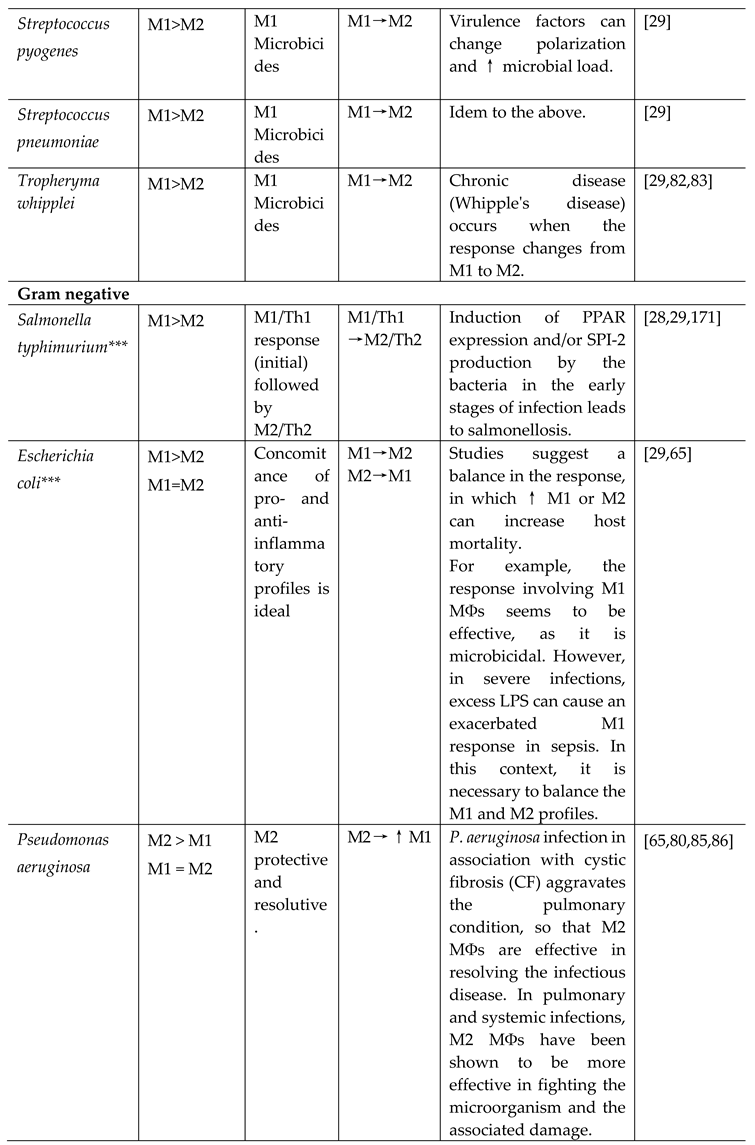         
|
*Considering chronologically the initial stage of the microenvironment of the microbicidal response and, taking into account that, in more advanced stages, the context of the repair microenvironment can alter the sub-population densities, thus, the numerical supremacy of a certain sub-population in the protective profile can be temporal, being more common at the beginning of the infection (microbicidal action) and tending to decrease toward the end (resolution and repair), therefore, not ruling out the importance of both profiles in the course of the infection, but only highlighting the protective profile in terms of prevention or attenuation of the disease outcome. Moreover, considering that macrophage polarization is only one part of a more complete and complex immune response involving other cells and effectors. As well as general host conditions. **As a form of microbial evasion and reversal of the protective response to acute stages. ***The presence of these commensals in the human microbiome causes a balance between M1 and M2 subpopulations under normal conditions, the Table, as well as the entire text, addresses contexts in which the microorganism is potentially pathogenic and/or when interaction with pathogenic strains occurs. Thus, this review does not address some interactions between polarization and commensals of the human microbiome (such as Enterococcus faecalis, Bacillus subtilis, or H. pylori). ****Ka et al. [10] propose that in this infection model, as well as in most acute (such as sepsis) and chronic infections, there is no polarization, but rather a response model in which MOs and MФs alter their repertoires in a kinetic function (early vs. late stages of infection), thus M1 MФs and M2 have significant impacts on infection and disease progression, as well as in cases for resolution of infection. *****An appropriate response to the vital importance of the infected organ.
4. Virulence factors of microorganisms modulating the polarization patterns of MФs
In the context of host-microorganism coevolution, the MФs present important strategies to neutralize microbial aggression while using others to preserve tissue integrity, considering a "double-edged sword" scenario of their opposite profiles (M1 and M2) [20]. On the other hand, as in the Spy versus Spy scenario, microorganisms can also present diverse virulence mechanisms (production of soluble molecules, alteration of surface antigens, biofilm formation, among others) to counterattack in order to survive the host's attack, becoming a commensal or favoring and amplifying the infectious focus. In this context, the immune system has a preferential polarization profile of its cells to orchestrate a microbicidal mechanism that can eradicate the infectious disease and promote tissue repair. Consequently, a change in this profile can favor the pathogen to the detriment of the host, as has been observed in infections by various microorganisms [28,29,33]. Of clinical relevance, while the host mounts an orchestrated specific response, differences between strains of the microorganism can be related to the most diverse virulence mechanisms, aimed at counteracting the host by modifying this response, as is extensively highlighted for the change from a Th1 to Th2 profile and vice versa, thus altering the main mechanism for eradicating the infectious disease.
As already mentioned, obligate or facultative intracellular pathogens that invade MФs during infection may find it easier to modulate polarization, favoring the survival of the microorganism and the establishment of the disease (e.g. Mtb, L. monocytogenes, HCMV). Xavier et al. [91] demonstrated that Brucella spp. can survive and multiply in MФs through the activation of PPARγ, which favors an M2/Th2-type response that is not protective for the host. Furthermore, the production of the Omp25 protein by the microorganism is reported to change the profile of the response to an M2 phenotype [29].
Mycobacterium tuberculosis has numerous virulence mechanisms to facilitate its survival in the context of infection. As discussed in the topic on programming, Fe sequestration is a metabolic mechanism performed by M1 MФs that limits the availability of this essential element to the microorganism. In the context of Mtb infections, Fe is also crucial for the growth of mycobacteria, so the host's protective response via M1 MФs provides this microbicidal mechanism that is synergistic with others, preventing active TB. However, a high production of siderophores (mycobactins) by Mtb can increase the bioavailability of Fe to Mtb, and this virulence mechanism is responsible for favoring mycobacterial growth [28]. In this sense, Kahnert et al. [107] showed that a programming of M2 MФs, activated/amplified by the profile involving intense IL-4 production, predisposes to active TB by favoring the intracellular persistence of Mtb due to a weaker response of M2 MФs. This M2 MФs/Th2 profile is achieved by some virulence factors of Mtb, for example, in which the microorganism manages to decrease nitrosative stress (decreases •NO concentration) induced by MФs, and/or increases its bioavailability to Fe (increase in bacterioferritin), actions which, respectively, decrease the mycobactericidal response of MФs and increase the conditions favorable to the multiplication of mycobacteria. Therefore, the authors concluded that, in an environment of timeless predominance of M2 MФs, the control that limits the proliferation of mycobacteria is lost, so that the intracellular persistence of Mtb favors active TB.
Other virulence factors of Mtb are related to its resistance to host defense. The CFP-10, ESAT-6 proteins and the secretion of the CFP10:ESAT6 complex by Mtb are able to inhibit the trans-activation of LPS, which induces the NF-ĸB factor, blocking the transcriptional profile of the M1 program, therefore inhibiting the production of proinflammatory cytokines, the production of iNOS and thus regulating the production of oxidants (mainly •NO) to concentrations below those essential, compromising the anti-mycobacterial response [29,182].
In addition, iNOS activity and consequent •NO formation, present in the functional repertoire of M1 MФs, is essential for the containment of some pathogens, especially Mtb [183]. With this target specifically established, it has been shown that Mtb and other MФs obligate intracellular pathogens such as Leishmania spp. are controlled by an M1/Th1-type host protective response, mainly due to iNOS activity. On the other hand, an increased microbial load has been reported in microenvironments where Arg1 activity prevails over iNOS (indicating a supremacy of M2-type phenotypic sub-populations), as is seen in lesions by Leishmania spp, in which parasites can modulate the polarization of M2 MФs through a virulence strategy that involves altering membrane markers that promote recognition only by these MФs [28,183].
The initial immune response to pathogens, like Mtb, involves the recognition of PAMPs by PRRs in defense cells, such as MФs, which subsequently leads to polarization. Shen et al. [31] showed that Mtb can elevate the expression of IRAK-M, a modulator with mechanisms associated with inhibition of phosphorylation and dissociation of IRAK-1 and -4 in down-stream signaling of the interaction between mycobacterial PAMP and the TLR of MФ, decreasing the activation of more down-stream mediators, such as NF-κB, IRF7 and JNK, and the associated Th1 response and M1-type polarization, therefore favoring Mtb intracellular survival and persistence of infection. Richardson et al. [110] showed that the response involving Mtb PAMP interaction with TLR-2 and the down-stream activation of ERK increases the concentration of IL-10 and decreases that of IL-12, which favors the polarization of M2 MФs and decreases the polarization toward M1, favoring the multiplication of mycobacteria and persistence of the infection. Bénard et al. [184] showed that B cells, in addition to producing antibodies, can produce type 1 IFN-γ and, when accumulated in the lungs of animals infected with Mtb, induce a polarization of M2 MФs, in a context involving STAT-1 signaling.
The literature highlights that human exposure to M. leprae does not generate a relevant proinflammatory immune response, partly because the mycobacterium promotes weak stimulation of the immune system in terms of activation of cells that release TNF-α. However, prior vaccination with BCG reinforces this response, protecting the host. In addition, M. leprae has virulence mechanisms that prevent the Th1 and M1 MФs program response, such as the presence of its phenolic glycolipid (PGL1) that alters polarization toward M2, with low levels of ILs 1β and 6, and TNF-α being observed, and favors the humoral response to the detriment of activation via T cells with activation of the Th1/M1 program [115].
Salmonella typhimurium can produce the mediator SPI-2 which inhibits the relocation of the NADPH oxidase complex (Nox2) in phagosomes, potentially decreasing the microbicidal activity of M1 MФs mediated by ROS and favoring the persistence of the disease [29]. Coxiella burnetii elicits an M1-type protective response, but an atypical M2-type response can favor chronic disease, with chronic evolution of fever and an increase in the associated lethality rate [29].
The chronic form of the disease caused by T. whipplei, in Whipple's disease, is associated with a change in the response pattern from the M1 MФs profile to a predominance of M2, forming spongy body-type tissue infiltrates. Desnues et al [82] demonstrated a predominant M2 MФs population profile for the chronic and systemic form of the disease, and for chronic intestinal disease [83]. In tularemia, an intense formation of cell debris and infiltrates in the lungs associated with spherocytosis may lead to a more expressive polarization of M2 MФs [96].
Unlike the virulence mechanisms discussed so far, in which microorganisms favor themselves by switching from an M1 to M2 MФs profile, some viruses benefit from switching from M2 to M1 (Table 4) [33,148]. Furthermore, the polarization of MФs against C. neoformans reveals a pleiotropic and multipolarization context, in the various phases of infection and in different microenvironments, in which both response profiles (M1 and M2) can alternate as protective, and this microorganism can use diverse virulence mechanisms to alter the polarization toward a profile that aggravates the disease [28,185].
Through its core protein (HCVc), HCV can manipulate the polarization of MФs [157]. Activation through HCVc binding to TLR-2 of MOs and MФs results in the induction of proinflammatory mediators (IRAK-1 kinase, NF-κB, MAPK and TNF-α) and subsequently, in the transition of the response, activation of the M2 profile occurs. However, the interaction of HCVc with TLR2/STATs also results in a impaired polarization in both profiles, M1 and M2, in circulating MOs and resident Kupffer cells, and decreases phagocytosis, leading to a dysfunction of the effective response and favoring the persistence of the infection. In addition, an induction of proliferation and hepatic recruitment of CD4+ T cells induced by M2 MФs is observed [157].
5. Clinical implications of polarization: perspectives for research, diagnosis, prognosis, and therapy of inflammatory and/or infectious diseases
Worldwide, various research groups have explored experimental in vivo (animal) and in vitro models of MФs polarization, underpinning the current understanding of polarization concepts, exploring experimental models of healing and repair (in general, presenting a programming impairment and population selection of M2-type polarization), or models of infection and acute inflammation (usually to promote M1-type polarization). However, even in these models, there can be variation, as seen here, where polarization depends on the stimulus and microenvironment. On the other hand, in vitro studies with MФs from human MOs isolated from the peripheral blood of supposedly "healthy" subjects can present marked disadvantages for a real understanding of polarization, since the presence of a neglected pathological state can commit the MOs (already in the bloodstream) to a certain functional program (priming toward M1 or M2). This implies that two samples isolated from the same patient at different times can yield different polarization profiles, if in one period the patient has an acute infection/inflammation that compromises the M1 program, and in another a repair/healing process or infection that compromises the MOs to an M2 response. The same applies to the study of polarization in specific experimental models of acute inflammation (predominantly M1 MФs) or repair and healing (predominantly M2 MФs) [28,29,30,31,32,33].
With regard to the diagnosis and prognosis of inflammatory/infectious diseases, the polarization profile of MФs seems to be a good indicator, since the levels of specific cytokines together with functional assessments of MФs and transcriptional profiles indicate the numerical prevalence of a given subpopulation. This topic seems more advanced in terms of non-infectious inflammatory diseases, with potential clinical value in the diagnosis of cancer and heart disease, in the first case, due to the specificity of the sub-populations found in certain types of tumor and, in the second, due to the fact that 10% of cells that are not cardiomyocytes in the heart are MФs [186,187,188]. Also, as another example of a diagnostic finding, CD4+ T cells activated by M2 MФs, massively guided by the HCVc of HCV, induce specific antibodies against the virus, which can be detected at different stages, including the most advanced ones, in diagnostic tests for HCV [157].
With regard to therapeutic applications, the possible development of vaccines or drugs could be supported by the immunomodulation of MФs, boosting or inhibiting the polarization programming of a given sub-population of MФs, according to what favors the protection of the host against the pathogen and/or repair/healing in an infectious process involving an exacerbated proinflammatory response, in this context, and may even have the reprogramming of already polarized MФs as a therapeutic target [39,71,148,189]. Support for the therapeutic control of MФs polarization in the management of patients with infectious diseases has been proposed [148]. However, more studies, including in vivo studies (i.e. randomized controlled trials, RCTs), should be conducted to gain a better understanding of the mechanisms underlying polarization and its real clinical significance.
With implications for preventive (vaccination) and therapeutic strategies, Quiding-Järbrink et al. [30] showed that prior vaccination against H. pylori favors the M1 MФs profile in infections by this microorganism, and that this profile can exacerbate chronic and severe gastritis and predispose to stomach cancer, since these MФs are found in abundance in this precancerous lesion. Furthermore, these findings may be relevant to diagnosis and therapy, in the first case by quantifying these MФs and in the second by modulating this polarization. The literature establishes a similar mechanism regarding BCG vaccination inducing M1 polarization in the protective profile against TB.
Based on the findings of Shen et al. [31], regarding the virulence mechanisms of Mtb and the control of the associated MФs polarization, the authors proposed considerations in future studies for the development of anti-Mtb vaccines, aiming to reinforce the host's immune response to the microorganism. Richardson et al. [110] also showed that signaling involving the interaction between Mtb PAMPs and TLR-2 could be an important pharmacological target, aiming in the clinic to reinforce the polarization of M1 MФs in order to contain the multiplication of mycobacteria and the persistence and progression of TB.
Macrophages are considered important therapeutic targets in P. jirovecii disease, according to Nandakumar et al. [124], aiming to modulate these cells and the context of the general immune response in the lung, with the aim of fully resolving the disease and avoiding damage and sequelae to the organ.
Ponce et al. [133] speculated on the purinergic system (ATP/adenosine) as a therapeutic target involving the polarization of MФs in the treatment of Chagas cardiomyopathy, observing promising results by inhibiting the CD73 enzyme, with significant improvements in the cardiac condition in animal models of T. cruzi infection.
With regard to the clinical-therapeutic considerations of MФs polarization in the context of HIV-1 infection, Gerngross et al. [177] showed that maintaining the polarization of MФs and microglia to an M2 profile serves as a reservoir of simian immunodeficiency virus (SIV) in an animal model of encephalitis, with control of such polarization being an attractive therapeutic target for treating complications of HIV-1 infection. Similar clinical-therapeutic implications for controlling the balance of MФs during HIV-1 infection were made by Williams et al. [178], when evaluating the impacts of the virus on these cells and the deleterious effect on the spleen in the long term. Representing the molecular diversity of polarization control, the CCR5 co-receptor, responsible for the tropism of HIV-1 by MФs, can alter the level of infection by the virus and the polarization of these cells, and is therefore considered a therapeutic target for generating resistance to HIV-1 infection [148]. Also, with regard to the redox balance in HIV-1 infection, Brundu et al. [190] showed, using a similar experimental animal model of infection and with extrapolations to HIV-1 infection, that depletion of cysteine and reduced glutathione (GSH) can occur in lymphoid organs in the early stages of infection, this being linked to an imbalance of polarization with a predominance of the Th2/M2 profile. To correct the redox status, the authors treated the animals with an N-acetyl-cysteine (NAC) supplement and, as a result, the polarization control was adjusted, with a decrease in ILs -4 and -5 and an increase in MФs, promoting less activation of M2 MФs. Not only limited to the search for new therapeutic alternatives, but also under the influence of therapies already implemented in clinical practice, Galvão-Lima et al [149] showed the influence of antiretroviral therapy in PL-HIV-1 on the functional profile of macrophages in response to different stimuli.
Given the context of persistent HCV infection, as already discussed here, Zhang et al. [157] speculated that therapeutic strategies involving the blocking of HCVc from the virus would be a viable alternative for treating infected patients, since the majority of those infected develop chronic hepatitis due to this protein, which is considered a virulence factor.
During clinical cases of sepsis and disseminated intravascular coagulation (DIC), the classic use of corticosteroids, causing immunosuppression and a change in polarization to the M2 profile, is useful. LPS, which is very abundant in E. coli, is one of the main causes of sepsis, a disorder highly characterized by systemic inflammation in disseminated infections. Aiming to find a therapeutic alternative to contain sepsis in microbial infections and improve the survival of those affected, Song et al. [191] tested a molecule derived from a marine fungus, Bis-N-Norgliovictine, and observed a decrease in the levels of immune mediators of inflammation and the polarization of M1 MФs and the damage associated with it, as well as greater survival in affected animals with a reduction in liver and lung damage.
Interestingly, the M2 MФs related to the resolution of the infectious focus is also related to the production of analgesic mediators to combat pain related to the inflammatory response and infectious disease, which raises the possibility of modulating this M2 profile as a potential therapeutic target in pain relief [17].
Nevertheless, therapeutic applications can be seen both in conventional pharmacological therapy, as shown in studies that address the influence of molecules and drugs already in use on polarization, and in more advanced fields, since genetic and epigenetic manipulation of polarization programming and reprogramming, with possible applications in the pharmaceutical field, has been explored [160]. In addition, representing a recent field in medicine and therapeutics, the use of lasers can modulate polarization [9], a strategy that has recently been employed as an adjuvant therapy in COVID-19 [192].
6. Final considerations
Accumulating evidence suggests that specific polarization patterns of MФs can be directed by intra- and extracellular pathogens that cause acute and chronic diseases. In some infections, due to the vital importance of the affected tissue/organ, the response pattern may be altered to protect the tissue and maintain its functionality. There may be a kinetic predominance of one subpopulation at any given time or a balance between the two or more. Therefore, the classical chronological sequence model with a predominance of M1 MФs subpopulations in the early stages of infection to initiate and maintain the inflammatory response and the arrival of M2 MФs in more advanced stages to resolve and promote repair/healing is not applicable to all infection models. There are polarization models that propose a more dynamic, temporal process with multiple ramifications in which more than two MФs subsets coexist.
Host as well as virulence factors of the microorganism can modulate polarization in addition to early pathogen-specific polarization characteristics. The control of MФs polarization can be considered an important therapeutic target for the implementation of preventive strategies (vaccines) and the treatment of infectious diseases. However, further studies should be conducted to gain a more in-depth knowledge of the specific underlying mechanisms of polarization and evidences of the clinical impact on humans (need for RCTs). Host factors, including genetics, comorbidities, and general health, are additional players that may modulate MФs polarization.
Finally, from research to diagnosis, prognosis and therapy of infectious diseases, the polarization of MФs has considerable clinical potential. The study of the drugs that are currently available and the search for new molecules that can modulate the polarization is also a field of current interest.
Author Contributions
C.D.C., conceptualization, design, bibliography analysis, writing-original draft preparation, writing-review and editing, visualization; M.R.P.L.B, visualization, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgements
This work is a complementary part of CDC's doctoral dissertation. This is the original title (doctoral dissertation, Thesis): “Influência da laserterapia sobre o burst oxidativo e perfil microbicida de macrófagos derivados de monócitos humanos” (English version: Influence of laser therapy on the oxidative burst and microbicidal profile of human monocyte-derived macrophages). This study was financed in part by the “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brasil”. Finance Code 001. CDC thanks CAPES for the scholarship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cerdeira, C.D. Complicações e sequelas neurológicas e psiquiátricas da COVID-19: uma revisão sistemática. Vittalle – Revista de Ciências da Saúde 2022, 34, 20–42. [Google Scholar] [CrossRef]
- Cerdeira, C.D.; Pereira de Araújo, M.; Jorge-Ferreira, C.B.R.; Dias, A.L.T.; Brigagão, M.R.P.L. Explorando os estresses oxidativo e nitrosativo contra fungos: um mecanismo subjacente à ação de tradicionais antifúngicos e um potencial novo alvo terapêutico na busca por indutores oriundos de fontes naturais. Rev. Colomb. Cienc. Quím. Farm. 2021, 50, 100–157. [Google Scholar] [CrossRef]
- Cerdeira, C.D.; Chavasco, J.K.; Brigagão, M.R.P.L. Tempol decreases the levels of reactive oxygen species in human neutrophils and impairs their response to Mycobacterium tuberculosis. Rev. Colomb. Cienc. Quím. Farm. 2022, 51, 123–151. [Google Scholar]
- Cerdeira, C.D.; da Silva, J.J.; Netto, M.F.R.; Boriollo, M.F.G.; Santos, G.B.; dos Reis, L.F.C.; Brigagão, M.R.P.L. Talinum paniculatum leaves with in vitro antimicrobial activity against reference and clinical strains of Staphylococcus aureus interfere with oxacillin action. Rev. Colomb. Cienc. Quím. Farm. 2020, 49, 432–451. [Google Scholar] [CrossRef]
- Amorim, C.F.; Souza, A.S.; Diniz, A.G.; Carvalho, N.B.; Santos, S.B.; Carvalho, E.M. Functional Activity of Monocytes and Macrophages in HTLV-1 Infected Subjects. Negl. Trop. Dis. 2014, 8, e339. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Saeed, A.F.U.H.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduction and Targeted Therapy 2023, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- León, B.; Ardavín, C. Monocyte-derived dendritic cells in innate and adaptive immunity. Immunology and Cell Biology 2008, 86, 320. [Google Scholar] [CrossRef] [PubMed]
- Koncz, G.; Jenei, V.; Tóth, M.; Váradi, E.; Kardos, B.; Bácsi, A.; Mázló, A. Damage-mediated macrophage polarization in sterile inflammation. Front. Immunol. 2023, 14, 1169560. [Google Scholar] [CrossRef] [PubMed]
- de Castro, M.S.; Miyazawa, M.; Nogueira, E.S.C.; Chavasco, J.K.; Brancaglion, G.A.; Cerdeira, C.D.; et al. Photobiomodulation enhances the Th1 immune response of human monocytes. Lasers in Medical Science 2022, 37, 135–148. [Google Scholar] [CrossRef]
- Ka, M.B.; Daumas, A.; Textoris, J.; Mege, J.L. Phenotypic diversity and emerging new tools to study macrophage activation in bacterial infectious diseases. Front. Immunol. 2014, 5, 500. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Reports 2014, 6, 13. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper T cell clone. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar] [CrossRef]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: the role of inflammation in Alzheimer disease. Nature, 2016, 16, 358–372. [Google Scholar] [CrossRef]
- Tatano, Y.; Shimizu, T.; Tomioka, H. Unique Macrophages Different from M1/M2 Macrophages Inhibit T Cell Mitogenesis while Upregulating Th17 Polarization. Scientific Reports 2014, 4, 4146. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Hill, A. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ma, L.; Deng, D.; Zhang, T.; Han, L.; Xu, F.; Huang, S.; Ding, Y.; Chen, X. M2 macrophage polarization: a potential target in pain relief. Front Immunol. 2023, 14, 1243149. [Google Scholar] [CrossRef]
- Chávez-Galán, L.; Olleros, M.L.; Vesin, D.; Garcia, I. Much more than M1 and M2 macrophages, there are also CD169+ and TCR+ macrophages. Front. Immunol. 2015, 6, 263. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: phenotypical vs. functional differentiation. Frontiers in Immunology 2014, 5, 514. [Google Scholar] [CrossRef]
- Lugo-Villarino, G.; Vérollet, C.; Maridonneau-Parini, I.; Neyrolles, O. Macrophage polarization: convergence point targeted by Mycobacterium tuberculosis and HIV. Frontiers in Immunology. 2011, 2, 10. [Google Scholar] [CrossRef]
- Huang, C.B.; Alimova, Y.; Ebersole, J.L. Macrophage polarization in response to oral commensals and pathogens. Pathogens and Disease 2016, 74, ftw011. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.F. Metchnikoff’s legacy in 2008. Nature Immunol 2008, 9, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. A polarizing question: do M1 and M2 microglia exist? Nature Neuroscience 2016, 19, 987–991. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Montenegro-Burke, J.F.; Sutton, J.A.; Rogers, L.M.; et al. Lipid profiling of polarized human monocyte-derived macrophages. Prostaglandins & other Lipid Mediators 2016, 127, 1–8. [Google Scholar] [CrossRef]
- Martinez, F.O.; Fernando, O.; Siamon, G.; Mantovani, A. Transcriptional profiling of the human monocyte- to-macrophage differentiation and polarization: new molecules and patterns of gene expression. Journal of Immunology (Baltimore, Md.:1950) 2006, 177, 7303–7311. [Google Scholar] [CrossRef]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nature Immunology 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Muraille, E.; Leo, O.; Moser, M. Th1/Th2 paradigm extended: macrophage polarization as an unappreciated pathogen-driven escape mechanism? Frontiers in Immunology 2014, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Benoit, M.; Desnues, B. ; Mege, J-L. Macrophage Polarization in Bacterial Infections. J Immunol, 2008; 181, 3733–3739. [Google Scholar]
- Quiding-Järbrink, M.; Raghavan, S.; Sundquist, M. Enhanced M1 Macrophage Polarization in Human Helicobacter pylori-Associated Atrophic Gastritis and in Vaccinated Mice. PLoS ONE 2010, 5, e15018. [Google Scholar] [CrossRef]
- Shen, P.; Li, Q.; Ma, J.; et al. IRAK-M alters the polarity of macrophages to facilitate the survival of Mycobacterium tuberculosis. BMC Microbiology 2017, 17, 185. [Google Scholar] [CrossRef]
- Das, A.; Sinha, M.; Datta, S.; et al. Monocyte and Macrophage Plasticity in Tissue Repair and Regeneration. The American Journal of Pathology 2015, 185. [Google Scholar] [CrossRef] [PubMed]
- Labonte, A.C.; Tosello-Trampont, A.-C.; Hahn, Y.S. The Role of Macrophage Polarization in Infectious and Inflammatory Diseases. Mol. Cells. 2014, 37, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Galván-Peña, S.; O’Neill, L.A.J. Metabolic reprograming in macrophage polarization. Frontiers in Immunology 2014, 5, 6. [Google Scholar] [CrossRef]
- Juhas, U.; Ryba-Stanisławowska, M.; Szargiej, P.; et al. Different pathways of macrophage activation and polarization. Postepy Hig. Med. Dosw. (online) 2015, 69, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.A. Arginine metabolism: boundaries of our knowledge. J Nutr 2007, Suppl 2, 1602S–1609S. [Google Scholar] [CrossRef]
- Mills, E.L.; O’Neill, L.A. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal. Eur. J. Immunol. 2016, 46, 13–21. [Google Scholar] [CrossRef]
- Kajahn, J.; Franz, S.; Rueckert, E.; et al. Artificial extracellular matrices composed of collagen I and high sulfated hyaluronan modulate monocyte to macrophage differentiation under conditions of sterile inflammation. Biomatter 2012, 2, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Ogle, M.E.; Segar, C.E.; Sridhar, S.; Botchwey, E.A. Monocytes and macrophages in tissue repair: Implications for immunoregenerative biomaterial design. Experimental Biology and Medicine 2016, 241, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Pizzolla, A.; Hultqvist, M.; Nilson, B.; et al. Reactive Oxygen Species Produced by the NOX2 Complex in Monocytes Protect Mice from Bacterial Infections. J. Immunol. 2012, 188, 5003–5011. [Google Scholar] [CrossRef]
- Bogdan, C.; Röllinghoff, M.; Diefenbach, A. Reactive oxygen and reactive nitrogen intermediates in innate and specific immunity. Current Opinion in Immunology 2000, 12, 64. [Google Scholar] [CrossRef]
- Wong, K.-W.; Jacobs Jr, W.R. Mycobacterium tuberculosis exploits human interferon γ to stimulate macrophage extracellular trap formation and necrosis. JID 2013, 208, 109. [Google Scholar] [CrossRef]
- de Oliveira-Junior, E.B.; Bustamante, J.; Newburger, P.E.; et al. The Human NADPH Oxidase: Primary and Secondary Defects Impairing the Respiratory Burst Function and the Microbicidal Ability of Phagocytes. Scandinavian Journal of Immunology 2011, 73, 420–427. [Google Scholar] [CrossRef]
- Takei, H.; Araki, A.; Watanabe, H.; Ichinose, A.; Sendo, F. Rapid killing of human neutrophils by the potent activator phorbol 12-myristate 13-acetate (PMA) accompanied by changes different from typical apoptosis or necrosis. J Leukoc Biol. 1996, 59, 229–240. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; et al. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532. [Google Scholar] [CrossRef] [PubMed]
- Röhm, M.; Grimm, M.J.; D'Auria, A.C.; et al. NADPH oxidase promotes neutrophil extracellular trap formation in pulmonary aspergillosis. Infect Immun. 2014, 82, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Stoiber, W.; Obermayer, A.; Steinbacher, P.; Krautgartner, W.D. The Role of Reactive Oxygen Species (ROS) in the Formation of Extracellular Traps (ETs) in Humans. Biomolecules 2015, 5, 702–723. [Google Scholar] [CrossRef]
- Chow, O.A.; Von Kockritz-Blickwede, M.; Bright, A.T.; Hensler, M.E.; Zinkernagel, A.S.; Cogen, A.L.; et al. Statins enhance formation of phagocyte extracellular traps. Cell Host Microbe 2010, 8, 445. [Google Scholar] [CrossRef] [PubMed]
- Bartneck, M.; Keul, H.A.; Zwadlo-Klarwasser, G.; Groll, J. Phagocytosis independent extracellular nanoparticle clearance by human immune cells. Nano Lett. 2010, 10, 59–63. [Google Scholar] [CrossRef]
- Aulik, N.A.; Hellenbrand, K.M.; Czuprynsk, C.J. Mannheimia haemolytica and Its Leukotoxin Cause Macrophage Extracellular Trap Formation by Bovine Macrophages. Infection and Immunity 2012, 80, 1923–1933. [Google Scholar] [CrossRef]
- Liu, P.; Wu, X.; Liao, C.; Liu, X.; Du, J.; et al. Escherichia coli and Candida albicans Induced Macrophage Extracellular Trap-Like Structures with Limited Microbicidal Activity. PLoS ONE 2014, 9, e90042. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Zou, X.-B.; Chai, Y.-F.; Yao, Y-M. Macrophage Polarization in Inflammatory Diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Boe, D.M.; Curtis, B.J.; Chen, M.M.; et al. Extracellular traps and macrophages: new roles for the versatile phagocyte. J Leukoc Biol. 2015, 97, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Doster, R.; Rogers, L.; Aronoff, D.; et al. Macrophages Produce Extracellular Traps in Response to Streptococcus agalactiae Infection. OFID 2015, 2 (Suppl 1), S235. [Google Scholar] [CrossRef]
- Je, S.; Quan, H.; Yoon, Y.; et al. Mycobacterium massiliense Induces Macrophage Extracellular Traps with Facilitating Bacterial Growth. PLoS ONE 2016, 11, e0155685. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, O.; Medina, E. The expanding world of extracellular traps: not only neutrophils but much more. Frontiers in Immunology 2013, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Espinosa, O.; Rojas-Espinosa, O.; Moreno-Altamirano, M.M.; et al. Metabolic requirements for neutrophil extracellular traps formation. Immunology 2015, 145, 213. [Google Scholar] [CrossRef] [PubMed]
- Van der Does, A.M.; Beekhuizen, H.; Ravensbergen, B. LL 37 directs macrophage differentiation toward macrophages with a proinflammatory signature. J. Immunol. 2010, 185, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Guimarães-Costa, A.B.; Rochael, N.C.; Oliveira, F.; et al. Neutrophil extracellular Traps reprogram IL-4/GM-CSF-induced Monocyte Differentiation to anti-inflammatory Macrophages. Frontiers in Immunology 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; et al. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to C. albicans. J Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef]
- Tan, H.-Y.; Wang, N.; Li, S.; et al. The Reactive Oxygen Species in Macrophage Polarization: Reflecting Its Dual Role in Progression and Treatment of Human Diseases. Oxidative Medicine and Cellular Longevity 2016, 2016, 2795090. [Google Scholar] [CrossRef]
- Neil,S. ; Huh, J.; Baronas, V.; et al. Oral administration of the nitroxide radical TEMPOL exhibits immunomodulatory and therapeutic properties in multiple sclerosis models. Brain, Behavior, and Immunity 2017, 62, 332–343. [Google Scholar] [CrossRef]
- Tillack, K.; Breiden, P.; Martin, R.; et al. T Lymphocyte Priming by Neutrophil Extracellular Traps Links Innate and Adaptive Immune Responses. The Journal of Immunology 2012, 20, 10. [Google Scholar] [CrossRef] [PubMed]
- Csillag, A.; Boldogh, I.; Pazmandi, K.; et al. , Pollen-Induced Oxidative Stress Influences Both Innate and Adaptive Immune Responses via Altering Dendritic Cell Functions. J. Immunol. 2010, 184, 2377–2385. [Google Scholar] [CrossRef] [PubMed]
- Mège, J.-L.; Mehraj, V.; Capo, C. Macrophage polarization and bacterial infections. Current Opinion in Infectious Diseases 2011, 24, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Dolgachev, V.A.; Yu, B.; Sun, L.; et al. IL-10 Overexpression Alters Survival in the Setting of Gram Negative Pneumonia Following Lung Contusion. Shock 2014, 41, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Munoz-Planillo, R.; Nunez, G. Sensing and reacting to microbes through the inflammasomes. Nat Immunol. 2012, 13, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Katz, B.P.; Spinola, S.M. Haemophilus ducreyi-Induced Interleukin-10 Promotes a Mixed M1 and M2 Activation Program in Human Macrophages. Infection and Immunity 2012, 80, 4426–4434. [Google Scholar] [CrossRef]
- Ogston, A. Report upon Micro-Organisms in Surgical Diseases. Br Med J. 1881, 1, 369. [Google Scholar] [CrossRef]
- Fournier, F.; Philpott, D.J. Recognition of Staphylococcus aureus by the innate immune system. Clinical Microbiology Reviews 2005, 18, 521. [Google Scholar] [CrossRef]
- Brown, A.F.; Leech, J.M.; Rogers, T.R.; McLoughlin, R.M. Staphylococcus aureus colonization: modulation of host immune response and impact on human vaccine design. Frontiers in Immunology 2014, 4, 1–20. [Google Scholar] [CrossRef]
- da Silva, J.J.; Cerdeira, C.D.; Chavasco, J.M.; et al. In vitro screening antibacterial activity of Bidens pilosa Linné and Annona crassiflora Mart. against oxacillin resistant Staphylococcus aureus (ORSA) from the aerial environment at the dental clinic. Rev Inst Med Trop Sao Paulo 2014, 56, 340. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Ma, H.-D.; Yin, X-Y.; et al. ; et al. Forkhead Box O1 Regulates Macrophage Polarization Following Staphylococcus aureus Infection: Experimental Murine Data and Review of the Literature. Clinic Rev Allerg Immunol. 2016, 51, 353–369. [Google Scholar] [CrossRef]
- Xu, F.; Kang, Y.; Zhang, H.; et al. Akt1-mediated regulation of macrophage polarization in a murine model of Staphylococcus aureus pulmonary infection. J. Infect. Dis. 2013, 208, 528–538. [Google Scholar] [CrossRef]
- Kasraie, S.; Niebuhr, M.; Kopfnagel, V.; et al. , Macrophages from patients with atopic dermatitis show a reduced CXCL10 expression in response to staphylococcal a-toxin. Allergy 2012, 67, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Prabhakara, R.; Harro, J.M.; Leid, J.G.; et al. Suppression of the Inflammatory Immune Response Prevents the Development of Chronic Biofilm Infection Due to Methicillin-Resistant Staphylococcus aureus. Infection and Immunity 2011, 79, 5010–5018. [Google Scholar] [CrossRef]
- Flannagan, R.S.; Heit, B.; Heinrichs, D.E. Antimicrobial Mechanisms of Macrophages and the Immune Evasion Strategies of Staphylococcus aureus. Pathogens 2015, 4, 826–868. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; et al. Staphylococcus aureus biofilms prevent macrophage phagocytosis and attenuate inflammation in vivo. J. Immunol. 2011, 186, 6585–6596. [Google Scholar] [CrossRef]
- Krysko, O.; Holtappels, G.; Zhang, N.; et al. Alternatively activated macrophages and impaired phagocytosis of S. aureus in chronic rhinosinusitis. Allergy 2011, 66, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Hanke, M.L.; Angle, A.; Kielian, T. MyD88-Dependent Signaling Influences Fibrosis and Alternative Macrophage Activation during Staphylococcus aureus Biofilm Infection. PLoS ONE 2012, 7, e42476. [Google Scholar] [CrossRef]
- Shaughnessy, L.M.; Swanson, J.A. The role of the activated macrophage in clearing Listeria monocytogenes infection. Front. Biosci 2007, 12, 2683–2692. [Google Scholar] [CrossRef]
- Desnues, B.; Raoult, D.J.; Mege, L. IL-16 is critical for Tropheryma whipplei replication in Whipple’s disease. J. Immunol. 2005, 175, 4575–4582. [Google Scholar] [CrossRef]
- Desnues, B.; Lepidi, H.; Raoult, D.J.; Mege, L. Whipple disease: intestinal infiltrating cells exhibit a transcriptional pattern of M2/alternatively activated macrophages. J. Infect. Dis. 2005, 192, 1642–1646. [Google Scholar] [CrossRef]
- Sica, A.; Erreni, M.; Allavena, P.; et al. Macrophage polarization in pathology. Cell. Mol. Life Sci. 2015, 72, 4111–4126. [Google Scholar] [CrossRef] [PubMed]
- Bielen, K.; ‘S Jongers, B.; Boddaert, J.; et al. Biofilm-Induced Type 2 Innate Immunity in a Cystic Fibrosis Model of Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2017, 7, 274. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.D.; McClellan, S.A.; Barrett, R.P.; et al. IL-33 shifts macrophage polarization, promoting resistance against Pseudomonas aeruginosa keratitis. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1524–1532. [Google Scholar] [CrossRef] [PubMed]
- Vicetti Miguel, R.D.; Harvey, S.A.K.; LaFramboise, W.A.; et al. Human Female Genital Tract Infection by the Obligate Intracellular Bacterium Chlamydia trachomatis Elicits Robust Type 2 Immunity. PLoS ONE 2013, 8, e58565. [Google Scholar] [CrossRef]
- Rottenberg, M.E.; Gigliotti-Rothfuchs, A.; Wigzell, H. The role of IFN-γ in the outcome of chlamydial infection. Curr. Opin. Immunol. 2002, 14, 444–451. [Google Scholar] [CrossRef]
- Jupelli, M.; Shimada, K.; Chiba, N.; et al. Chlamydia pneumoniae Infection in Mice Induces Chronic Lung Inflammation, iBALT Formation, and Fibrosis. PLoS ONE 2013, 8, e77447. [Google Scholar] [CrossRef]
- Cesta, M.C.; Zippoli, M.; Marsiglia, C.; Gavioli, E.M.; Cremonesi, G.; Khan, A.; Mantelli, F.; Allegretti, M.; Balk, R. Neutrophil activation and neutrophil extracellular traps (NETs) in COVID-19 ARDS and immunothrombosis. Eur J Immunol. 2023, 53, e2250010. [Google Scholar] [CrossRef]
- Xavier, M.N.; Winter, M.G.; Spees, A.M.; et al. PPARγ-mediated increase in glucose availability sustains chronic Brucella abortus infection in alternatively activated macrophages. Cell Host Microbe 2013, 14, 159–170. [Google Scholar] [CrossRef]
- Liu, P.; Lin, Y.-W.; Burton, F.H.; Wei, L.-N. M1-M2 balancing act in white adipose tissue browning – a new role for RIP140. Adipocyte 2015, 4, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.-H.; Xu, Y.; Chen, X.-M.; et al. , Macrophage cell death upon intracellular bacterial infection. Macrophage (Houst) 2015, 26, e779. [Google Scholar] [CrossRef]
- Paciello, I.; Silipo, A.; Lembo-Fazio, L.; et al. Intracellular Shigella remodels its LPS to dampen the innate immune recognition and evade inflammasome activation. Proc. Natl. Acad. Sci. USA 2013, 110, E4345–E4354. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, V.; Textoris, J.; Amara, A.B.; et al. Monocyte Responses in the Context of Q Fever: From a Static Polarized Model to a Kinetic Model of Activation. JID 2013, 208, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Mares, C.A.; Sharma, J.; Li, Q.; et al. Defect in efferocytosis leads to alternative activation of macrophages in Francisella infections. Immunology and Cell Biology 2011, 89, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.P.; Verriere, T.; Assim, M.; et al. Heme Oxygenase-1 Dysregulates Macrophage Polarization and the Immune Response to Helicobacter pylori. J. Immunol. 2014, 193, 3013–3022. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Sharma, P.K.; Mukhopadhaya, A. Vibrio cholerae porin OmpU mediates M1-polarization of macrophages/monocytes via TLR1/TLR2 activation. Immunobiology 2015, 220, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xia, J.; Li, T.; et al. Shp2 Deficiency Impairs the Inflammatory Response Against Haemophilus influenzae by Regulating Macrophage Polarization. JID 2016, 214, 625–633. [Google Scholar] [CrossRef]
- Lasky, E.C.; Olson, R.M.; Brown, C.R. Macrophage Polarization during Murine Lyme Borreliosis. Infect. Immun. 2015, 83, 2627–2635. [Google Scholar] [CrossRef]
- Radolf, J.D.; Arndt, L.L.; Akins, D.R.; et al. Treponema pallidum and Borrelia burgdorferi lipoproteins and synthetic lipopeptides activate monocytes/macrophages. J. Immunol. 1995, 154, 2866–2877. [Google Scholar] [CrossRef]
- Seiler, K.P.; Vavrin, Z.; Eichwald, E.; et al. Nitric oxide production during murine Lyme disease: lack of involvement in host resistance or pathology. Infect. Immun. 1995, 63, 3886–3895. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.P.; Zachary, J.F.; Teuscher, C.; et al. Dual role of interleukin-10 in murine Lyme disease: regulation of arthritis severity and host defense. Infect. Immun. 1999, 67, 5142–5150. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Zhang, N.; Wooten, R.M. Borrelia burgdorferi elicited-IL-10 suppresses the production of inflammatory mediators, phagocytosis, and expression of co-stimulatory receptors by murine macrophages and/or dendritic cells. PLoS ONE 2013, 8, e84980. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.C.; Lefimil, C.; Rodas, P.I.; et al. Neisseria gonorrhoeae Modulates Immunity by Polarizing Human Macrophages to a M2 Profile. PLoS ONE, 2015, 10, e0130713. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, G.; Shaik-Dasthagirisaheb, Y.B.; Huang, N.; et al. Immunologic environment influences macrophage response to Porphyromonas gingivalis. Molecular Oral Microbiology, 2017, 32, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Kahnert, A.; Seiler, P.; Stein, M.; et al. Alternative activation deprives macrophages of a coordinated defense program to Mycobacterium tuberculosis. Eur. J. Immunol. 2006, 36, 631–647. [Google Scholar] [CrossRef]
- Brasil. Dados sobre a Turbeculose no Brasil. 2023.
- Forrellad, M.A.; Klepp, L.I.; Gioffré, A.; et al. Virulence factors of the Mycobacterium tuberculosis complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef] [PubMed]
- Richardson, E.T.; Shukla, S.; Sweet, D.R.; et al. Toll-like receptor 2-dependent extracellular signal-regulated kinase signaling in Mycobacterium tuberculosis-infected macrophages drives antiinflammatory responses and inhibits Th1 polarization of responding T cells. Infect. Immun. 2015, 83, 2242–2254. [Google Scholar] [CrossRef]
- Roy, S.; Guler, R.; Parihar, S.P.; et al. Batf2/Irf1 Induces Inflammatory Responses in Classically Activated Macrophages, Lipopolysaccharides, and Mycobacterial Infection. The Journal of Immunology 2015, 194, 6035–6044. [Google Scholar] [CrossRef]
- Schreiber, T.; Ehlers, S.; Heitmann, L.; et al. Autocrine IL-10 induces hallmarks of alternative activation in macrophages and suppresses antituberculosis effector mechanisms without compromising T cell immunity. J. Immunol. 2009, 183, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, M.V.; Brooks, M.N.; Morris, J.D.; et al. Mycobacterium tuberculosis activates human macrophage peroxisome proliferator-activated receptor gamma linking mannose receptor recognition to regulation of immune responses. J. Immunol. 2010, 185, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Aira, N.; Andersson, A.-M.; Singh, S.K.; et al. Species dependent impact of helminth-derived antigens on human macrophages infected with Mycobacterium tuberculosis: Direct effect on the innate antimycobacterial response. PLoS Negl. Trop. Dis. 2017, 11, e0005390. [Google Scholar] [CrossRef] [PubMed]
- Fallows, D.; Peixoto, B.; Kaplan, G.; et al. Mycobacterium leprae alters classical activation of human monocytes in vitro. Journal of Inflammation 2016, 13, 8. [Google Scholar] [CrossRef]
- da Silva, B.J.; Barbosa, M.G.M.; Andrade, P.R.; et al. Autophagy Is an Innate Mechanism Associated with Leprosy Polarization. PLoS Pathog. 2017, 13, e1006103. [CrossRef]
- da Silva, T.P.; Bittencourt, T.L.; de Oliveira, A.L.; et al. Macrophage Polarization in Leprosy–HIV Co-infected Patients. Front. Immunol. 2020, 11, 1493. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-F.; Hong, Y.-X.; Feng, G-J.; et al. Lipopolysaccharide-Induced M2 to M1 Macrophage Transformation for IL-12p70 Production Is Blocked by Candida albicans Mediated Up-Regulation of EBI3 Expression. PLoS ONE 2013, 8, e63967. [Google Scholar] [CrossRef]
- Richardson, J.P.; Moyes, D.L. Adaptive immune responses to Candida albicans infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Cerdeira, C.D.; da Silva, J.J.; Netto, M.F.R.; et al. Talinum paniculatum: a plant with antifungal potential mitigates fluconazole-induced oxidative damage-mediated growth inhibition of Candida albicans. Rev. Colomb. Cienc. Quím. Farm. 2020, 49, 401–431. [Google Scholar] [CrossRef]
- Cerdeira, C.D.; Brigagão, M.R.P.L.; Carli, M.L.; et al. Low-level laser therapy stimulates the oxidative burst in human neutrophils and increases their fungicidal capacity. J. Biophotonics 2016, 9, 1180. [Google Scholar] [CrossRef]
- Lefèvre, L.; Authier, H.; Stein, S.; et al. LRH-1 mediates anti-inflammatory and antifungal phenotype of IL-13-activated macrophages through the PPARγ ligand synthesis. Nat. Commun. 2015, 6, 6801. [Google Scholar] [CrossRef] [PubMed]
- Reales-Calderón, J.A.; Aguilera-Montilla, N.; Corbí, A.L.; et al. Proteomic characterization of human proinflammatory M1 and anti-inflammatory M2 macrophages and their response to Candida albicans. Proteomics 2014, 14, 1503–1518. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, V.; Hebrink, D.; Jenson, P.; et al. Differential Macrophage Polarization from Pneumocystis in Immunocompetent and Immunosuppressed Hosts: Potential Adjunctive Therapy during Pneumonia. Infect Immun. 2017, 85, e00939–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. , Wang, F.; Bhan, U.; et al. TLR9 Signaling Is Required for Generation of the Adaptive Immune Protection in Cryptococcus neoformans-Infected Lungs. The American Journal of Pathology 2010, 177, 754–765. [Google Scholar] [CrossRef]
- Eastman, A.J.; He, X.; Qiu, Y.; et al. Cryptococcal HSP70 homologue Ssa1 contributes to pulmonar expansion of C. neoformans during the afferent phase of the immune response by promoting macrophage M2 polarization. J. Immunol. 2015, 194, 5999–6010. [Google Scholar] [CrossRef] [PubMed]
- Roos, D. Chronic granulomatous disease. Br Med Bull 2016, 118, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.S.; Oliveira, A.F.; da Silva, T.A.; et al. Paracoccin Induces M1 Polarization of Macrophages via Interaction with TLR4. Frontiers in Microbiology 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- da Silva, T.A.; Roque-Barreira, M.C.; Casadevall, A.; Almeida, F. Extracellular vesicles from Paracoccidioides brasiliensis induced M1 polarization in vitro. Scientific Reports 2016, 6, 35867. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.M.; Scott, A.L. Antecedent Nippostrongylus infection alters the lung immune response to Plasmodium berghei. Parasite Immunol. 2017, 39, e12441. [Google Scholar] [CrossRef]
- Guirelli, P.M.; Angeloni, M.B.; Barbosa, B.F.; et al. Trophoblast-macrophage crosstalk on human extravillous under Toxoplasma gondii infection. Placenta 2015, 36, 1106e1114. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Q.; Chao, J.; et al. Polarization of macrophages induced by Toxoplasma gondii and its impact on abnormal pregnancy in rats. Acta Trop. 2015, 143, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ponce, N.E.; Sanmarco, L.M.; Eberhardt, N.; et al. CD73 Inhibition Shifts Cardiac Macrophage Polarization toward a Microbicidal Phenotype and Ameliorates the Outcome of Experimental Chagas Cardiomyopathy. The Journal of Immunology, 2016, 197, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.F.; Cerdeira, C.D. Eventos tromboembólicos associados à cardiopatia chagásica: revisão de literature. Revista da Universidade Vale do Rio Verde, Três Corações 2015, 13, 619–629. [Google Scholar] [CrossRef]
- Cabalén, M.E.; Cabral, M.F.; Sanmarco, L.M.; et al. Chronic Trypanosoma cruzi infection potentiates adipose tissue macrophage polarization toward an anti-inflammatory M2 phenotype and contributes to diabetes progression in a diet induced obesity model. Oncotarget 2016, 7, 13400–13415. [Google Scholar] [CrossRef] [PubMed]
- McCartney Francis, N.; Jin, W.; Belkaid, Y.; et al. Aberrant host defense against Leishmania major in the absence of SLPI. J. Leukoc. Biol. 2014, 96, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Saldarriaga, O.A.; Spratt, H.; et al. Transcriptional Profiling in Experimental Visceral Leishmaniasis Reveals a Broad Splenic Inflammatory Environment that Conditions Macrophages toward a Disease-Promoting Phenotype. PLoS Pathog. 2017, 13, e1006165. [Google Scholar] [CrossRef] [PubMed]
- Rani, R.; Jordan, M.B.; Divanovic, S.; et al. IFN-γ–Driven IDO Production from Macrophages Protects IL-4R–Deficient Mice against Lethality during Schistosoma mansoni Infection. The American Journal of Pathology 2012, 180, 2001–2008. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, H.; Chen, L.; et al. Schistosoma japonicum infection induces macrophage polarization. The Journal of Biomedical Research 2014, 28, 299–308. [Google Scholar] [CrossRef]
- dos Reis, L.F.C.; Cerdeira, C.D.; Gagliano, G.S.; et al. Alternate-day fasting, a high-sucrose/caloric diet and praziquantel treatment influence biochemical and behavioral parameters during Schistosoma mansoni infection in male BALB/c mice. Experimental Parasitology 2022, 240, 108316. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, P.; Chen, Y.; et al. Inhibition of Notch Signaling Attenuates Schistosomiasis Hepatic Fibrosis via Blocking Macrophage M2 Polarization. PLoS ONE, 2016, 11, e0166808. [Google Scholar] [CrossRef]
- Peon, A.N.; Espinoza-Jimenez, A.; Terrazas, L.I. Immunoregulation by Taenia crassiceps and its antigens. Biomed. Res. Int. 2013, 2013, 498583. [Google Scholar] [CrossRef]
- Zawistowska-Deniziak, A.; Basałaj, K.; Strojny, B.; et al. New Data on human Macrophages Polarization by Hymenolepis diminuta Tapeworm—an In Vitro study. Front. Immunol. 2017, 8, 148. [Google Scholar] [CrossRef]
- Tundup, S.; Srivastava, L.; Harn, D.A., Jr. Polarization of host immune responses by helminth-expressed glycans. Ann. N.Y. Acad. Sci. 2012, 1253, E1–E13. [Google Scholar] [CrossRef] [PubMed]
- Tawill, S.; Le Goff, L.; Ali, F. Both free-living and parasitic nematodes induce a characteristic Th2 response that is dependent on the presence of intact glycans. Infect. Immun. 2004, 72, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.F.; Cerdeira, C.D.; Prado, A.C.; Dias, R.P.C.S.; Silva, R.B.V.; Vertêlo, P.C.; Silvério, A.C.P. ; Profile of people living with HIV at a reference center in contagious and infectious diseases in Belo Horizonte (MG, Brazil). Revista de Medicina e Saúde de Brasília 2020, 9, 76–89. [Google Scholar]
- Brasil. Boletim Epidemiológico AIDS e DST. Brasília: Ministério da Saúde, Secretaria de Vigilância em Saúde, PN de DST e AIDS, 2017, 48.
- Sang, Y.; Miller, L.C.; Blecha, F. Macrophage Polarization in Virus-Host Interactions. J Clin Cell Immunol. 2015, 6. [Google Scholar] [CrossRef]
- Galvão-Lima, L.J.; Espíndola, M.S.; Soares, L.S.; et al. Classical and alternative macrophages have impaired function during acute and chronic HIV-1 infection. Braz. J. Infect. Dis. 2017, 21, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Cassetta, L.; Rizzi, C.; Alfano, M.; Poli, G. M1 and M2a polarization of human monocyte-derived macrophages inhibits HIV-1 replication by distinct mechanisms. J Immunol. 2009, 182, 6237–6246. [Google Scholar] [CrossRef]
- Vertêlo, P.C.; Miranda, A.B.; Labanca, L.; Borges-Starling, A.L.; Cerdeira, C.D.; et al. Prevalence of VIH-1/HTLV-1 co-infection and behavioral risk among people living with HIV in Belo Horizonte, Brazil. Revista de Salud Pública 2022, 28, 75–85. [Google Scholar]
- Pedral-Sampaio, D.B.; Martins Netto, E.; Pedrosa, C.; et al. Co-Infection of tuberculosis and HIV/HTLV retroviruses: frequency and prognosis among patients admitted in a Brazilian hospital. Braz. J. Infect. Dis. 1997, 1, 31–35. [Google Scholar]
- Porto, A.F.; Neva, F.A.; Bittencourt, H. HTLV-1 decreases Th2 type of immune response in patients with strongyloidiasis. Parasite Immunol 2001, 23, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Marinho, J.; Galvão-Castro, B.; Rodrigues, L.C.; et al. Increased risk of tuberculosis with human T lymphotropic virus I infection: A case-control study. J. Acquir. Immune Defic. Syndr. 2005, 40, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Porto, A.F.; Santos, S.B.; Alcantara, L.; et al. HTLV-1 modifies the clinical and immunological response to schistosomiasis. Clin. Exp. Immunol. 2004, 137, 424–429. [Google Scholar] [CrossRef]
- Bastos, M.L.; Santos, S.B.; Souza, A.; et al. Influence of HTLV-1 on the clinical, microbiologic and immunologic presentation of tuberculosis. BMC Infect. Dis. 2012, 12, 199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.; Zhai, N.; et al. HCV core protein inhibits polarization and activity of both M1 and M2 macrophages through the TLR2 signaling pathway. Scientific Reports 2016, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Sun, Y.; Pu, J.; et al. Comparative virus replication and host innate responses in human cells infected with three prevalent clades (2.3.4, 2.3.2, and 7) of highly pathogenic avian influenza H5N1 viruses. J Virol. 2014, 88, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Toapanta, F.R.; Shirey, K.A.; et al. Potential role for alternatively activated macrophages in the secondary bacterial infection during recovery from influenza. Immunol Lett. 2012, 141, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.; Morris, P.; Dickman, M.J.; et al. The therapeutic potential of epigenetic manipulation during infectious diseases. Pharmacology & Therapeutics 2016, 167, 85–99. [Google Scholar] [CrossRef]
- Lian, Q.; Zhang, K.; Zhang, Z.; et al. Differential effects of macrophages subtypes on SARS-CoV-2 infection in a human pluripotent stem cell-derived model. Nature Communications 2022, 13, 2028. [Google Scholar] [CrossRef]
- da Silva Amorim, A.F.; Jesus Coimbra, M.; Podestá, M.H.M.C.; da Silva, A.O.; Cerdeira, C.D.; et al. Critical Factors Associated with Morbimortality in COVID-19 Patients Attended at a Brazilian Public Hospital: a Cross-Sectional Study. Journal of Health Sciences 2023, 25, 96–107. [Google Scholar] [CrossRef]
- Proctor, P. Tempol treatment of COVID-19. Free Radic Biol Med. 2020, 159, S95. [Google Scholar] [CrossRef]
- Brodin, P.; Davis, M.M. Human immune system variation. Nat Rev Immunol. 2017, 17, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Jaume, M.; Yip, M.S.; Cheung, C.Y.; et al. Anti-severe acute respiratory syndrome coronavirus spike antibodies trigger infection of human immune cells via a pH- and cysteine protease-independent FcγR pathway. J. Virol. 2011, 85, 10582–10597. [Google Scholar] [CrossRef] [PubMed]
- Yip, M.S.; Leung, N.H.; Cheung, C.Y.; et al. Antibody-dependent infection of human macrophages by severe acute respiratory syndrome coronavirus. Virol. J. 2014, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhao, J.; Van Rooijen, N.; et al. Evasion by stealth: inefficient immune activation underlies poor T cell response and severe disease in SARS-CoV-infected mice. PLoS Pathog. 2009, 5, e1000636. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Dong, C.; Xiong, S. IL-33 enhances macrophage M2 polarization and protects mice from CVB3-induced viral myocarditis. Journal of Molecular and Cellular Cardiology 2017, 103, 22–30. [Google Scholar] [CrossRef]
- Chan, G.; Bivins-Smith, E.R.; Smith, M.S. NF-κB and Phosphatidylinositol 3-Kinase Activity Mediates the HCMV-Induced Atypical M1/M2 Polarization of Monocytes. Virus Res. 2009, 144, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-S.; Tseng, Y.-H.; Chen, Y-C.; et al. M2 macrophage subset decrement is an indicator of bleeding tendency in pediatric dengue disease. Journal of Microbiology, Immunology and Infection 2018, 51, 829e838. [Google Scholar] [CrossRef]
- Eisele, N.A.; Ruby, T.; Jacobson, A.; et al. Salmonella require the fatty acid regulator PPAR for the establishment of a metabolic environment essential for long-term persistence. Cell Host Microbe 2013, 14, 171–182. [Google Scholar] [CrossRef]
- Almeida, P.E.; Roque, N.R.; Magalhães, K.G.; et al. Differential TLR2 downstream signaling regulates lipid metabolism and cytokine production triggered by Mycobacterium bovis BCG infection. Biochim Biophys Acta 2014, 1841, 97–107. [Google Scholar] [CrossRef]
- Gomes, M.S.; Boelaert, J.R.; Appelberg, R. Role of iron in experimental Mycobacterium avium infection. J Clin Virol. 2001, 20, 117–122. [Google Scholar] [CrossRef]
- Weinberg, J.B.; Volkheimer, A.D.; Rubach, M.P. Monocyte polarization in children with falciparum malaria: relationship to nitric oxide insufficiency and disease severity. Scientific Reports 2016, 6, 29151. [Google Scholar] [CrossRef]
- Stijlemans, B.; Guilliams, M.; Raes, G.; et al. African trypanosomosis: From immune escape and immunopathology to immune intervention. Veterinary Parasitology 2007, 148, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Stempin, C.C.; Motrán, C.C.; Aoki, M.P.; et al. PD-L2 negatively regulates Th1-mediated immunopathology during Fasciola hepatica infection. Oncotarget 2016, 7, 77721–77731. [Google Scholar] [CrossRef]
- Gerngross, L.; Lehmicke, G.; Belkadi, A.; et al. Role for cFMS in maintaining alternative macrophage polarization in SIV infection: implications for HIV neuropathogenesis. Journal of Neuroinflammation 2015, 12, 58. [Google Scholar] [CrossRef]
- Williams, D.W.; Engle, E.L.; Shirk, E.N.; et al. Splenic Damage during SIV Infection Role of T-Cell Depletion and Macrophage Polarization and Infection. The American Journal of Pathology 2016, 186, 2068–2087. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.J.; Maury, W. The role of mononuclear phagocytes in Ebola virus infection. J Leukoc Biol. 2018, 104, 717–727. [Google Scholar] [CrossRef]
- Rogers, K.J.; Brunton, B.; Mallinger, L.; Bohan, D.; Sevcik, K.M.; Chen, J.; et al. IL-4/IL-13 polarization of macrophages enhances Ebola virus glycoprotein-dependent infection. PLoS Negl Trop Dis 2019, 13, e0007819. [Google Scholar] [CrossRef] [PubMed]
- Stantchev, T.S.; Zack-Taylor, A.; Mattson, N.; et al. Cytokine Effects on the Entry of Filovirus Envelope Pseudotyped Virus-Like Particles into Primary Human Macrophages. Viruses 2019, 11, 889. [Google Scholar] [CrossRef]
- Ganguly, N.; Giang, P.H.; Gupta, C.; Basu, C.K.; Siddiqui, I.; Salunke, D.M.; Sharma, P. Mycobacterium tuberculosis secretory proteins CFP-10, ESAT-6 and the CFP10:ESAT6 complex inhibit lipopolysaccharide-induced NF-kB transactivation by downregulation of reactive oxidative species (ROS) production. Immunology and Cell Biology 2008, 86, 98–106. [Google Scholar] [CrossRef]
- Singh, R.; Manjunatha, U.; Boshoff, H.I.; Há, Y.H.; Niyomrattanakit, P.; Ledwidge, R.; Dowd, C.S.; Lee, I.Y.; Kim, P.; Zhang, L.; Kang, S.; Keller, T.H.; Jiricek, J.; Barry, C. E 3rd. PA-824 Kills Nonreplicating Mycobacterium tuberculosis by Intracellular NO Release. Science 2008, 322, 1392–1395. [Google Scholar] [CrossRef]
- Bénard, A.; Sakwa, I.; Schierloh, P.; et al. B Cells Producing Type I Interferon Modulate Macrophage Polarization in Tuberculosis. American Journal of Respiratory and Critical Care Medicine. [CrossRef]
- Davis, M.J.; Tsang, T.M.; Qiu, Y.; et al. Macrophage M1/M2 polarization dynamically adapts to changes in cytokine microenvironments in Cryptococcus neoformans infection. mBio 2013, 4, e00264–13. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.-F.; Wang, H. Monocyte and macrophage differentiation: circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomarker Research 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Kalish, S.V.; Lyamina, S.V.; Usanova, E.A.; et al. Macrophages Reprogrammed In Vitro Towards the M1 Phenotype and Activated with LPS Extend Lifespan of Mice with Ehrlich Ascites Carcinoma. Med Sci Monit Basic Res. 2015, 21, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Nahrendorf, M.; Swirski, F.K. Abandoning M1/M2 for a Network Model of Macrophage Function. Circ Res. 2016, 119, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Lyamina, S.V.; Kruglov, S.V.; Vedenikin, T.Y.; et al. Alternative reprogramming of M1/M2 phenotype of mouse peritoneal macrophages in vitro with interferon γ and interleukin 4. Bull Exp Biol Med. 2012, 152, 548–51. [Google Scholar] [CrossRef]
- Brundu, S.; Palma, L.; Picceri, G.G.; et al. Glutathione Depletion Is Linked with Th2 Polarization in Mice with a Retrovirus-Induced Immunodeficiency Syndrome, Murine AIDS: Role of Proglutathione Molecules as Immunotherapeutics. J. Virol. 2016, 90, 7118–7130. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Dou, H.; Gong, W.; et al. Bis N norgliovictin, a small molecule compound from marine fungus, inhibits LPS induced inflammation in macrophages and improves survival in sepsis. Eur. J. Pharmacol. 2013, 705, 49–60. [Google Scholar] [CrossRef]
- de Matos, B.T.L.; Buchaim, D.V.; Pomini, K.T.; et al. Photobiomodulation therapy as a possible new approach in COVID-19: a systematic review. Life (Basel) 2021, 11, 580. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



