Submitted:
28 March 2023
Posted:
30 March 2023
You are already at the latest version
Abstract
Carbapenems are considered for treating Klebsiella pneumoniae and other Enterobacteriaceae infections, especially if they are not susceptible to other generally prescribed antibiotics, i.e., if they show resistance. In such cases, antibiotic activity decreases, and most patients succumb to the infection. A better understanding of the disease pattern and resistance mechanisms could be gained by magnifying the genes that confer resistance to antibiotics. Therefore, studying the genes that confer resistance to carbapenems and any other antibiotics for that matter is indispensable for coming up with improved treatment options. This study included the analyses of co-resistance patterns between resistance genes-between drug classes and within the carbapenem-resistant genes, genomic context analysis of highly expressed carbapenem-resistant genes, and phylogenetic study of OXA-producing genes, plasmid incompatibility identification, and sequence type identification using MLST. The presence of ESBLs, MBLs, and SBLs across the downloaded genomes was studied. SHV-producing genes were found to co-occur with most of the resistant genes belonging to different drug classes. The plasmid incompatibility type IncFIB was found to be common among the highly expressed genes, and most of these genes were flanked by different families of insertion sequence (IS) elements. MLST study suggested that the presence of sequence types ST-11, ST-14, and ST-147 was common in the downloaded set of genomes.
Keywords:
Carbapenem
; Antimicrobial Resistance
; Klebsiella pneumoniae
; genomic context
Introduction and Literature Review
Resistance to antibiotics is a growing concern and a threat to human health. Carbapenems are generally considered last-line agents to treat antibacterial-resistant infections1. Annually, approximately 10 million deaths worldwide are predicted due to antibacterial-resistant infections2. Carbapenem-resistant Enterobacteriaceae (CRE), especially Carbapenem-resistant Klebsiella pneumoniae (CRKP), pose a serious problem to clinicians due to its increasing prevalence in hospital settings.
These pathogens are acquired nosocomially, especially in Intensive Care Unit (ICU) patients. If the prescribed antibiotic course is not completed or if it is overprescribed, the patient may develop CRKP infections.
Figure 1.
AMR in 2050, Band et al.
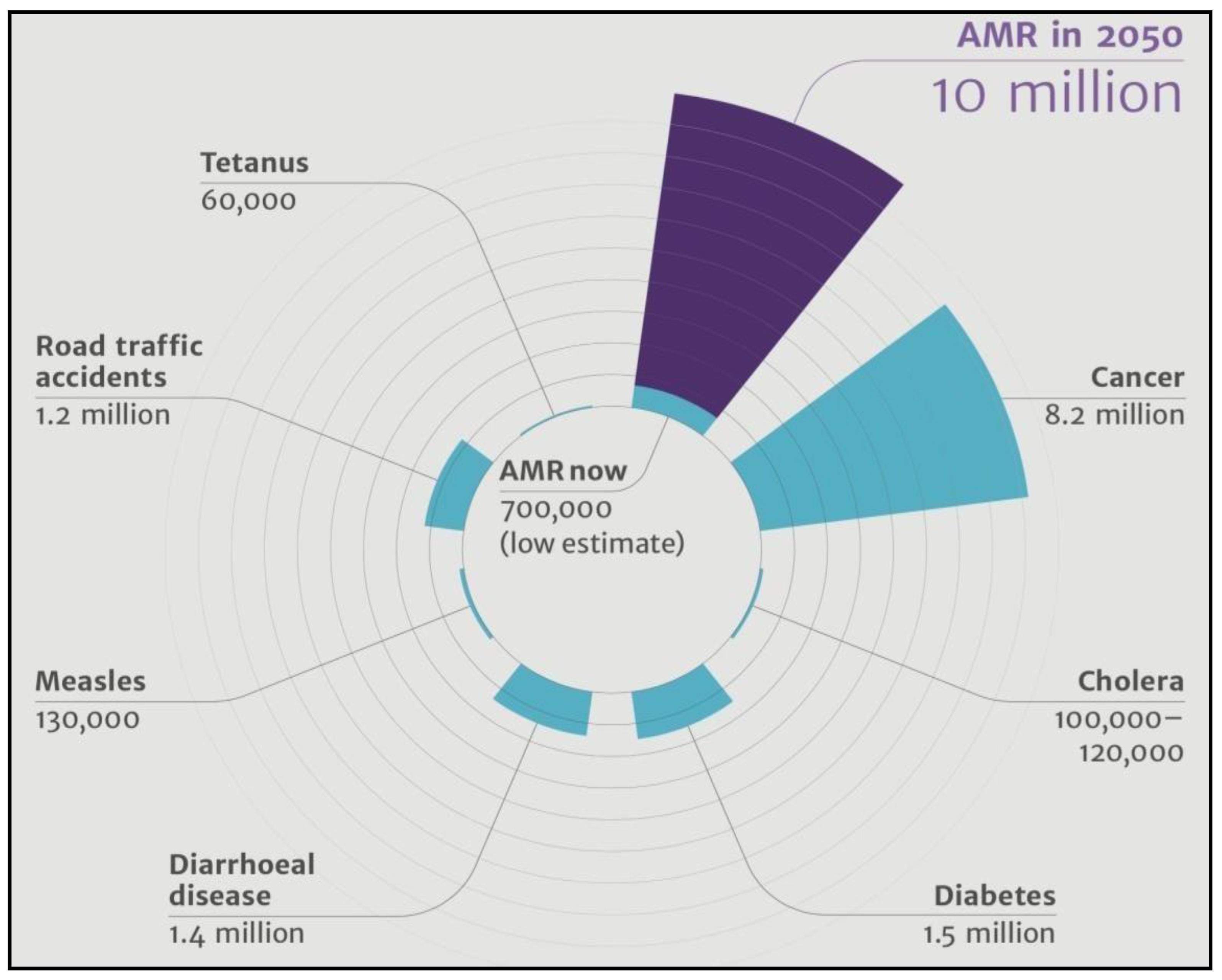
In the past, there were a lot of anomalies in coming up with a standard definition for antimicrobial-resistant resistant phenotypes. Consensus terms were proposed by Magiorakos et al. in 2012, which are Multiple Drug Resistance (MDR), extreme Drug Resistance (XDR), and Pan-drug Resistance (PDR). All currently encountered carbapenem-producing Enterobacteriaceae come under MDR and a substantial set under XDR. Generally used terms to denote Enterobacteriaceae that show resistance to carbapenems are Carbapenemase-producing carbapenem-resistant Enterobacteriaceae (Cp-CRE), Carbapenem-resistant Organisms (CRO), carbapenem-producing Organisms (CPO). Pathogens that are acquired nosocomially pose a serious threat to human health. The most common nosocomially acquired pathogens belong to the ESKAPE organisms3. The acronym ESKAPE pathogens exhibit MDR with high virulence. ESKAPE pathogens are Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumanii, Pseudomonas aeruginosa, and Enterobacter spp4.
The hunt for β-lactamase inhibitors began in the late 1960s5 when bacterial β- lactamases developed and threatened the use of penicillin. The first β-lactamase inhibitors were identified in 1976, and they were olivanic acids, which are natural products of the Gram-positive bacterium Streptomyces clavuligerus. Olivanic acids are broad-spectrum β-lactams with a "carbapenem backbone."
Two β-lactamase inhibitors6 were developed then,
- (i)
- Clavulanic acid from Streptomyces clavuligerus, which was the first clinically available inhibitor,
- (ii)
- Thienamycin from Streptomyces cattleya, which became the parent compound for all carbapenems.
Figure 2.
Thienamycin, olivanic acid, and clavulanic acid structure, Krisztina M. Papp- Wallace et al.
Figure 2.
Thienamycin, olivanic acid, and clavulanic acid structure, Krisztina M. Papp- Wallace et al.
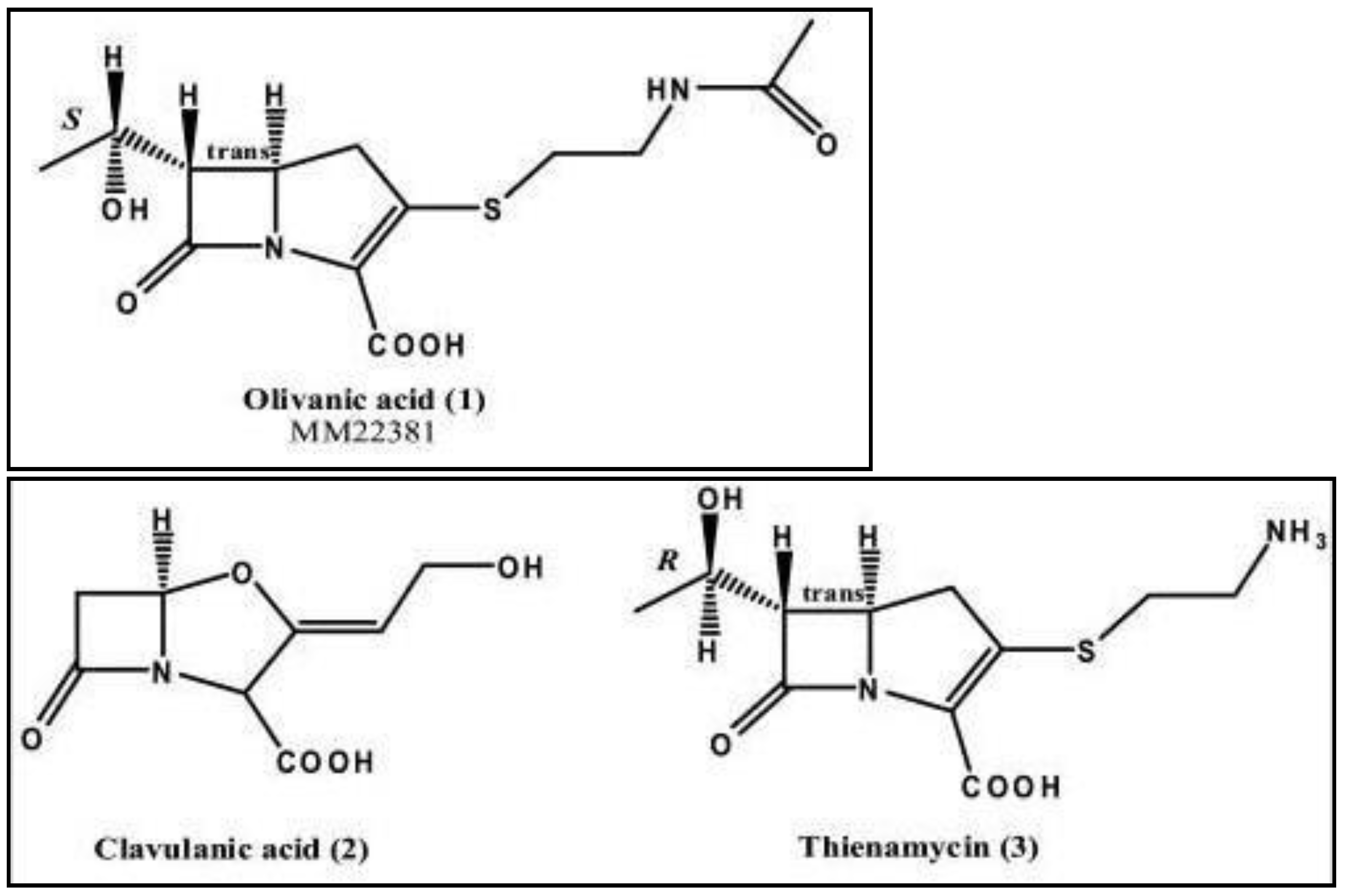
Although thienamycin was a natural product, after purification, the yields were low. It also exhibited instability in aqueous solutions, sensitivity to hydrolysis by mild bases, and high reactivity to nucleophiles, which all prompted a better alternative7. Therefore, its synthetic preparation gained importance, and this gave rise to imipenem.
Imipenem, an N-formimidoyl derivative8, and panipenem, a closely related carbapenem that was identified a little later, were found to be less sensitive to base hydrolysis and more stable than thienamycin. Imipenem became available in 1985, exhibiting great stability against β-lactamases and a high affinity for Penicillin- binding proteins (PBPs). Imipenem and panipenem later became susceptible to dihydropeptidase-1 (DHP-1) mediated deactivation in the human renal brush borders9. Therefore, inhibitors like cilastatin and betamipron were used for coadministration10. Carbapenems exhibiting more stability were later introduced, and some of the compounds that are currently in use include ertapenem, doripenem, meropenem, and biapenem and are also found to be stable towards hydrolysis by DHP-16.
Some important considerations include,
- (i)
- imipenem and doripenem are more potent than meropenem against Acinetobacter baumanii11,
- (ii)
- low Minimum inhibitory Concentration (MIC) of doripenem than imipenem and doripenem against A.baumanii and P.aeruginosa, and doripenem are least susceptible to hydrolysis by carbapenemases12,
- (iii)
- limited spectrum of ertapenem against P.aeruginosa11,
- (iv)
- MDR Mycobacterium tuberculosis can be treated with meropenem in combination with clavulanic acid13.
Klebsiella pneumoniae, when found in their natural intestinal residence, are harmless. But if found anywhere else in the body, they could turn into superbugs which could render them undefeatable by antibiotics. Some of the diseases caused by the bacterium are meningitis, urinary tract infections (UTIs), endocarditis, etc. Since certain strains of the Enterobacteriaceae family harbor genes that encode Extended Spectrum β-Lactamases (ESBLs), misuse of antibiotics could lead to bacterial resistance to carbapenems. The mechanisms by which bacterial strains mediate resistance are porin modifications or loss (ampC overexpression), efflux pumps, and hydrolyzing enzymes.
Carbapenems present a broad spectrum of antimicrobial activity. The presence of carbapenem (4:5 fused ring lactam) with a β-lactam ring, a double bond between C-1 and C-2, and sulfur at the C-1 structure give them more stability even against ESBLs. They inhibit the cell wall formation in bacteria by binding to the penicillin-binding proteins (PBP)14. For treating pan-drug-resistant bacteria, dual or synergistic therapy like carbapenem (doripenem) with colistin is considered15. This helps in fighting bacteria that are resistant to carbapenems but susceptible to colistin.
Carbapenems, like other β-lactams, do not pass through the cell wall of the bacteria16. They enter the Gram-negative bacteria through Outer Membrane Porin (OMP). Once they are inside the bacteria, they acylate the PBPs, which include transpeptidases, carboxypeptidases, and transglycosylase, and they help in the formation catalysis of peptidoglycan in the cell wall. Cell wall formation is a dynamic process where cell wall formation and autolysis occur at the same time, and when the PBPs are acylated, the autolysis continues, peptidoglycan weakens, and due to osmotic pressure, the cell bursts. The ability of carbapenems to bud to multiple different PBPs confers efficacy to this antibiotic (Hashizume et al.).
Certain enzymes called β-lactamases hydrolyze most β-lactam antibiotics, including carbapenems. According to the Ambler classification, β-lactamases can be classified as follows, class A (K. pneumoniae carbapenemase, KPC), class B Metallo-β- lactamases, MBL (New Delhi Metallo-β-lactamases, NDM) and class D (OXA-48- like carbapenemases)17. Class A carbapenemases contain serine at their active site that facilitates ring opening. Thus, they are called serine β-Lactamases (SBLs). KPC-2 and KPC-3 (Klebsiella pneumoniae Carbapenemases) are the most frequently reported of all variants. The β-lactam inhibitors of SBL are clavulanic acid, sulbactam, and tazobactam18. Class B β-lactamases contain two zinc cations at their active site and are classified into three groups: B1, B2, and B3. Thus, they are called Metallo β-Lactamases (MBLs)19. NDM family genes are an example of this class. Inhibitors are dipicolinic acid, EDTA, etc20. Class D β-lactamases contain serine at their active site, and they do not have any inhibitors. An example of this class of β-lactamases in the OXA-48 family genes21.
Figure 3.
Classification of different mechanisms of drug resistance in CRE, Suay-García et al.
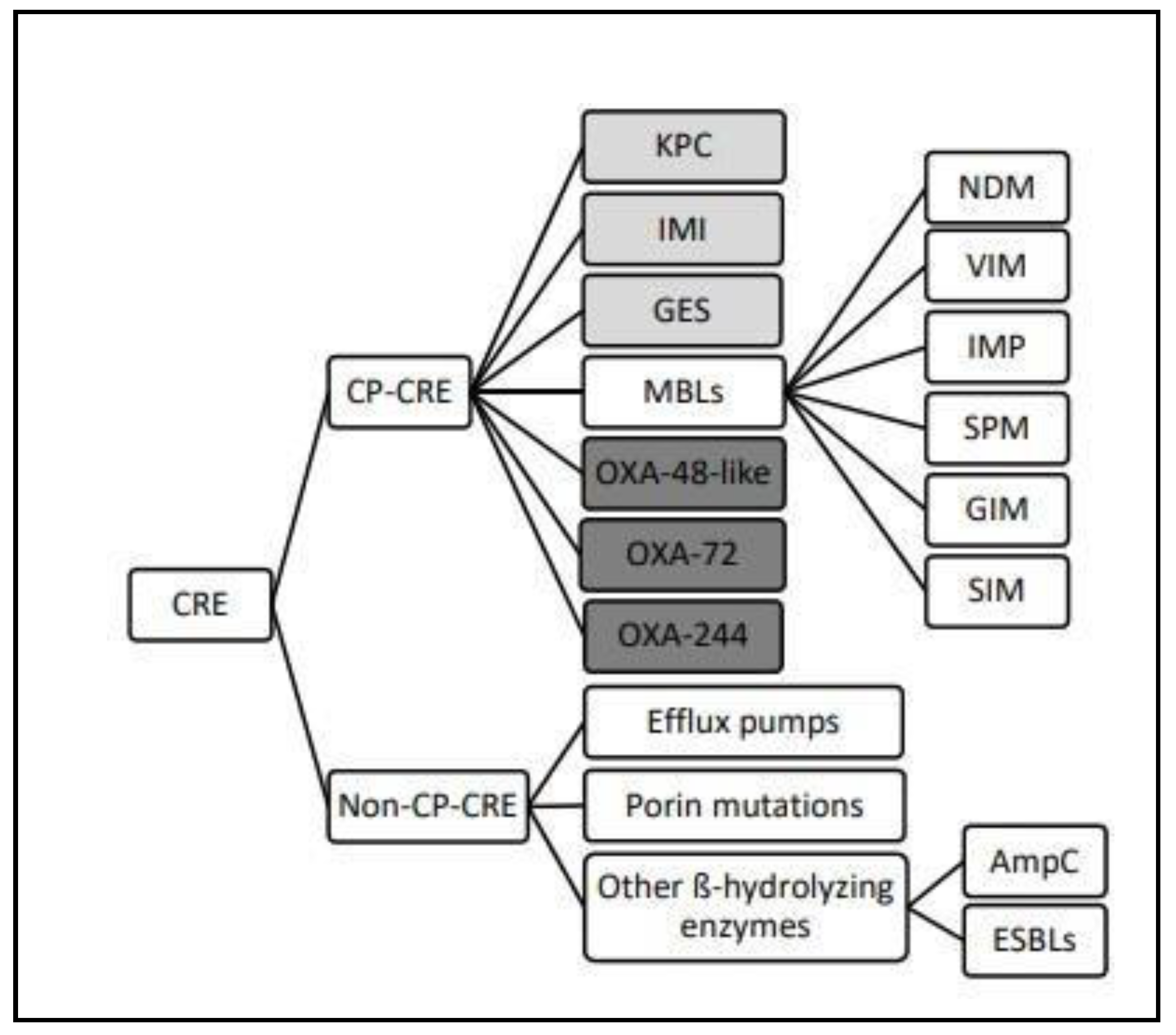
A study conducted by Xiaolin Yu et al. highlights that pathogens that harbor KPC-2 show resistance to all β-lactams and β-lactamase inhibitors like ceftazidime/avibactam, suggesting limited treatment options leading to a high rate of mortality. Apart from that, NDM has become a serious public threat because of the global dissemination of NDM-carrying pathogens and their presence on mobile genetic elements (MGEs) across different species of bacteria.
It has been shown that the alleles are usually carried on plasmids or MGEs, and the plasmid has a toxin-antitoxin system that prevents plasmid loss even in the absence of selective pressure caused by the antibiotic22. The most prevalent carbapenemase genes include blaKPC, blaNDM, blaOXA-48, and blaVIM. Clonal expansion is suggested to be responsible for the infection outbreaks; plasmid-mediated transfer is hard to detect and may be underestimated in studies23. The most common enzymatic cause of carbapenems is because of the KPC enzymes. blaKPC-2 and blaKPC-3 are the most common and frequently reported genes that come under the blaKPC class. In isolation, OXA-48-like carbapenemases bring about a weak hydrolysis of penicillin and carbapenems but not cephalosporins. This leads to difficulty in detection, and for this reason, they are called "the phantom menace," according to Poirel et al.
Because of the presence of carbapenem resistance genes in pathogenic strains, it has been declared a "global sentient event." Prolonged hospital stay and severe underlying illness are the main risk factors for the isolation of ESBL-producing organisms. This is associated with adverse clinical and economic outcomes like the increased length of stay and mortality rate, delay in the administration of effective therapy, and increased cost of care. According to Kwak et al., previous exposure to cephalosporins and carbapenems is the major cause of acquiring CRKP infections, while exposure to fluoroquinolones is protective. In a study by Flagas et al., 53 patients with CRKP isolation were compared with matched controls, and it was found that prior infection to antipseudomonal penicillins and fluoroquinolones were independent risk factors for CRKP infections24.
On a global scale, the prevalence of carbapenemase genes was studied, and it was observed that the KPC-producing Enterobacteriaceae are found to be endemic to countries like the United States, Argentina, and Colombia, Italy, and Greece. MBL NDM-1 is the most prevalent in countries like India, Sri Lanka, and Pakistan25. According to Walsh et al., India is the main reservoir of NDM-producing bacteria. OXA-48-like enzyme producers are seen mainly in the middle eastern countries and North Africa25.
As suggested by the guidelines of the European Committee on Antimicrobial Susceptibility Testing (EUCAST) and the Clinical and Laboratory Standard Institute (CLSI), carbapenems continue to be administered for Enterobacteriaceae infections. Due to the unavailability of doripenem in most countries, it has been removed from the EUCAST 2019 guidelines25. High virulence in K.pneumoniae helps them in invading the host's immune protective mechanisms and infect. High viscosity is noted along with high virulence in such strains but is not an essential factor. The carbapenem antibiotics that are used for treatment not only inhibit the formation of cell walls but also function by inhibiting β-lactamases and protein synthesis (DeJonge et al. & Karaiskos et al.).
Studying the carbapenem-resistant genes gives an idea about the genes that confer resistance against different classes of antibiotics. This study aims at understanding the co-resistance patterns, identifying the plasmid type using Plasmid Database (PLSDB) and sequence type identification using Multiple Locus Sequence Typing (MLST), and studying the upstream and downstream genes of the gene under study.
Materials and Methods
For studying the genomic context of carbapenem-resistant Klebsiella pneumoniae and analyzing the genes involved in resistance acquisition, 1025 complete, chromosome level assembled genomes were downloaded from the NCBI genome database. This required the installation of the Miniconda Package Manager to ensure that all bioinformatics tools used were up to date and functional. The main packages used for this study are ncbi-genome-download and Resistance Gene Identifier (RGI).
Downloading protein genome sequence from NCBI
A conda environment was created, and the ncbi-genome-download tool was used to download the protein genome sequences from the NCBI database (RefSeq).
Identification of AMR genes
The genomes that were downloaded were analyzed to detect the presence of antimicrobial resistance genes using RGI. It uses the Comprehensive Antibiotic Resistance Database (CARD) to identify the presence of AMR genes. RGI reads the ORFs using Prodigal if nucleotide fasta is submitted to predict the protein sequences26. Protein fasta files were used with the default blast search strategy, which led to the generation of a csv file. Genomes and their respective genes that confer resistance to different drug classes were obtained. The json files obtained as an output of blast (DIAMOND BLAST) search were used to generate a heatmap. A shell script was created that included '-t' (input viz protein), '-p' (algorithm viz DIAMOND BLAST), and '-n' (number of CPU threads-10). The codes used for the generation of the files are,

A heatmap in the form of a csv file was generated that showed different drug classes and their respective resistant gene.
Co-occurrence study
Co-occurrence of resistance genes was conducted between carbapenem genes and genes of other drug classes like aminoglycosides, fosfomycin, colistin, etc., and also within the carbapenem-resistant gene classes. The input for this study was the csv file that was generated as an output from the RGI heatmap. The first sheet of M.S. Excel consisted of the drug classes, and a copy was created in the second sheet. The individual carbapenem resistance gene classes in the second sheet were selected and filled (black fill) in order to find out the genes that conferred resistance along with the selected genes. A script or a code was run through macros that carried out the computation. The code (Figure 4) was written in such a way that the computation can be carried out only in the second sheet. Initially, the input (csv) file gave the perfect, strict, and no hits as 2, 1, and 0, respectively. These numbers were used for studying co-resistance patterns among different drug classes. The percentage of co-occurrence was calculated, and radar charts were created for better visualization of the data.
Genomic context study
For analyzing the genomic context of selected genes, a software called GeneSpy was used. It is a powerful visualization tool to study functional links between genes27. A database was built by providing the NCBI ftp links of the genomes. Using the ftp links, a MySQL-based database with the gfm files were built. The gene names were given as input, and a search was performed to build the genetic context. The neighboring genes and their assembly in either chromosome or plasmid are the output that was exported as a pdf file. The conserved gene context was manually analyzed from the results.
Phylogenetic analysis
Out of the 14 gene classes that were obtained as a result of the RGI-CARD analysis, a phylogenetic study was carried out for the OXA-48-like family of genes (8 genes). To conduct this analysis, software called Mega (Molecular Evolutionary Genetic Analysis) v.1128 was installed. The protein fasta files of the OXA-48-like genes were downloaded as a batch from NCBI Batch Entrez. The database was set to protein, and the accession numbers were provided. The fasta files were retrieved and were manually copied/pasted in the Mega software for the alignment of the sequences. Alignment was carried out with the help of ClustalW. A Maximum Likelihood tree was constructed with 1000 bootstraps. For better visualization of the tree, a Newick format of the tree was created and was submitted to the iTOL29 (Interactive Tree of Life) online web tool. A circular tree was generated.
PLSDB
Plasmid Database (PLSDB) is an online database for plasmids gathered from NCBI and INSDC30. Since resistance in microbes is spread through plasmids and other MGEs, this analysis is of importance to characterize the infection-causing pathogens. The metadata includes incompatibility type, date, and geographical location. The Plasmid Finder pipeline determines the incompatibility type. Plasmid types of CMY- 2, CTX-M-27, IMP-4, KPC-3, OXA-1, NDM-1, SHV-11, and VIM-1 producing genes were determined.
MLST
Sequence Type identification of the RefSeq genomes was done using the tseemann/mlst downloaded from the git repository. To get the results of MLST as a csv file, a bash script was run. Data was generated by the RGI and the pubMLST pipeline. The data from CARD live included 10,030 genomes, out of which 1025 were that of K.pneumoniae. A python script (Figure 5) was run in the directory having the json files and was used to parse the data from the downloaded json files. Analysis of data was carried out using M.S. Excel. A bar chart indicating the Sequence Type (ST) and the number of genomes was made.
Results
Carbapenem-resistant genes that were highly expressed were analyzed. They are CMY-2, CTX-M-27, IMP-4, NDM-1, OXA-1, KPC-3, VIM-1, and SHV-11.
Carbapenem-resistance gene detection
Using RGI-CARD software, the genes conferring resistance against the carbapenem class of antibiotics and against other antibiotics were identified. A total of 82 carbapenem-resistance genes were found among 1022 genomes. Among them, 43 genes coding for ESBLs, CMY-2, CTX-M-27, PER-1, PER-7, and 39 coded for SHV. Two genes that code for efflux pumps, i.e., KpnG and KpnH, two genes that code for an outer membrane protein porin, OmpK37, and OmpA, one suspected transcriptional activator of genes involved in multiple antibiotic resistance, and marA, were found. An lptD gene that is involved in lipopolysaccharide (LPS) assembly at the outer membrane surface and an SFO-1 gene were identified. Thirty-two genes that encode carbapenemases like NDM, KPC, OXA, IMP, and VIM were detected. The individual gene classes are listed below (Table 1).
The distribution of the carbapenem resistance genes among 1025 K.pneumoniae genomes was studied. Among the genomes, NDM expression was found to be the highest with 36%, followed by OXA-48 like family genes with 21% expression (Figure 6).
Co-occurrence analysis
Two co-occurrence studies were conducted, one with the selected set of carbapenem resistance genes against the resistance-conferring genes of all other drug classes and the other between the selected set of carbapenem resistance genes against other genes belonging to the same class. Genes occurring ≥ 60% are listed in the following sections.
Between drug classes
Co-occurrence of the following carbapenem resistance gene classes with respect to the other drug classes was observed.
KPC
Resistance genes of aminoglycosides, carbapenems, diaminopyrimidine, disinfecting agents and intercalating dyes, fluoroquinolones, fosfomycin, penem, monobactam, phenicol, rifamycin, tetracycline, and triclosan were found to be co-occurring with KPC-producing genomes. A total of 23 genes occurring ≥60% were recorded (Table 2). Eighty-one percent of genomes producing blaKPC were also found to harbor fosA3, a fosfomycin resistant gene, and 80.165% of genomes also harbored blaSHV-134.
NDM
Co-occurrence of 3 genes conferring resistance to aminoglycosides, diaminopyrimidine, and glycopeptides with respect to the NDM class of genes was observed (Table 3). Ninety-two percent of the genomes that produced blaNDM were also found to possess aph(3')-VI, which confer resistance to aminoglycosides.
OXA
Twenty-eight genes were found to be co-occurring with the OXA-48-like family genes. blaOXA harboring genomes were also found to possess genes that conferred resistance to aminoglycosides, carbapenems, cephalosporins, cephamycins, diaminopyrimidine, fluoroquinolones, lincosamide, macrolides, penam, nucleoside, rifamycin, and tetracyclines (Table 4). The highest co-occurrence pattern noted was with aph(3")-VI, which confers resistance to aminoglycosides with a co-occurrence rate of 96.42%.
Individual studies were conducted for the CMY and IMP classes of genes, but significant co-resistance patterns were not observed.
Within carbapenem drug class
Co-occurrence analysis between the resistance genes of the carbapenem drug class was carried out, and the results are listed below.
Genomes that harbored KPC were also found to possess SHV-134 (80.1%), SHV-7 (75%), SHV-30 (66.6%), SHV-42 (66.6%), and CMY-2 (60%) (Figure 7).
All OXA harboring genomes possessed NDM-3, NDM-4, and NDM-19. Additionally, 83.3% of them had SHV-110, 66.6% had SHV-30, IMP-4, and 61.8% had NDM-1 (Figure 8).
Individual studies were also conducted for the VIM, CMY, NDM, and IMP against the other carbapenem-resistance genes, but significant co-resistance patterns were not observed.
Genomic context analysis
A genomic context study was performed using software called GeneSpy to understand the inherent genetic structure. Five upstream genes and five genes downstream of the gene under study were visualized and analyzed. The conserved genetic context of each gene under study is represented.
CMY-2
The presence of Class C beta-lactamase CMY-2 was seen in 11 genomes and four genomes (GCF_019317065.1, GCF_019317185.1, GCF_019317265.1, GCF_019334705.1) had CMY-2 family Class C beta-lactamase that harbored blaCMY-2. The upstream genes were IS1380-like element ISEcp1 family transposase, a hypothetical protein, traV (type 4 transfer system lipoprotein), traB (conjugal transfer), another hypothetical protein, and traL (type 4 conjugative transfer system). The downstream genes were lipocalin family proteins, sugE (quarternary ammonia compound efflux SMR transporter protein), Ig-like domain-containing protein, a Dsbc family protein, and traC (type 4 secretion system protein) (Figure 9).
CTX-M-27
CTX-M-27 genes were observed in 11 genomes. The upstream genes wereIS1380- like element ISEcp1 family transposase, malate/lactate/ureidoglycolate dehydrogenase gene, an fdhf (formate dehydrogenase subunit α), an M20 family metallopeptidase, and a yitz MFS transporter. Also, a multidrug efflux transporter mdtM gene was observed. In the genome GCF_019334625.1, tetracycline antibiotic transporter tetR and tetA (Figure 10) were observed. The downstream genes wereISKra4 family transposase, a recombinase family protein, a qnrS1 (Quinolone-resistant pentapeptide repeat), IS3, and a serine hydrolase (Figure 11).
IMP-4
Nine genomes were found to harbor the IMP-4 gene. The upstream genes were recombinase family protein, a TN3 family transposase, an IS110/IS4321 family transposase, and a DUF6531 domain-containing protein. The downstream genes were qacG2 (ammonia compound efflux SMR transporter), aminoglycoside N- acetyltransferase aac(6')-Ib4, catB3 chloramphenicol O-acetyl transferase, NAD+-- rifamycin ADP-ribosyltransferase Arr-3, and a qacE (ammonia SMR transporter). Macrolide-resistant MFS transporter mrxA was found extremely downstream (Figure 12).
KPC-3
KPC-3 genes were found in 57 genomes. The upstream genes were istB, istA, tnpA (transposase), tnpR (resolvase), and a hypothetical protein. The downstream genes were IS1182/ISKpn6 family transposase, a ycfP (alpha/beta hydrolase), a nagZ (β-N- acetyl hexosaminidase), a thiK (thiamine kinase), and a lpoB (PBP-activator) (Figure 13).
NDM-1
Seventy-nine genomes were found to possess the NDM-1 producing genes. The upstream genes wereIS630/ISEc33 family transposase, IS30/ISAba125, APH(3')-VI family aminoglycoside O-phosphotransferase, IS3, repM, and ABC-F type protection protein MSr(E). The downstream genes were Ble-MBL, PRA1, protein disulfide reductase, DsbD N-terminal domain-containing protein, divalent cation tolerance protein CutA, and co-chaperones groES and groEL (Figure 14).
OXA-1
OXA-1 was found in 297 genomes. The upstream genes were fluoroquinolone acetylating gene, aminoglycoside 6’-N-acetyltransferase AAC(6’-Ib-Cr5), IS6/IS26 family transposase, Tn3/TnAs1 family transposase, and a tetR tetracycline-resistant transcription repressor and a tetA efflux MFS transporter. The downstream genes were CatB-related O-acetyltransferase, IS6/IS26 family transposase, a recombinase family protein, intl1 class 1 integron integrase, and a dfrA14 trimethoprim resistant dihydrofolate reductase (Figure 15). In Klebsiella pneumoniae WP_201507552.1 (GCF_904863295.1), blaOXA-1 was observed, and upstream to it, blaCTX was also observed (Figure 16).
SHV-11
Five hundred and sixty-two genomes containing SHV-11 were noted. The upstream genes were AAA family ATPase, a yitz, a bgaA-β galactosidase, Lac1 family DNA- binding transcriptional regulator, and a GNAT family N-acetyl transferase. The downstream genes were DeoR/GIpR family DNA-binding transcriptional regulator, NADP-dependent oxidoreductase, aldolase, and an HPr family phospho carrier (Figure 17).
VIM-1
VIM-1 was found to be present in 14 genomes. The upstream genes were intl1 class 1 integron integrase, IS6/IS26 family transposase, DeoR/GIpR family DNA-binding transcriptional regulator, class A β-lactamase SHV-200 (plasmid), and a hypothetical protein. urtB urea ABC transporter permease subunit and urtC genes were also observed. The downstream genes were aminoglycoside 6’-N-acetyltransferase AAC(6')-Ib4, aminoglycoside O-phosphotransferase APH(3'), ANT(3') family aminoglycoside nucleotidyltransferase, type B-2 chloramphenicol O-acetyltransferase CatB2, and qacE. mer family of genes and sulfonamide resistant dihydropteroate synthase sul1 was also observed (Figure 18)
Phylogenetic analysis
ClustalW alignment of the OXA-class genes of multiple species, which included Enterobacteriaceae and Proteobacteria, was carried out using the Mega software, and a maximum likelihood tree was constructed. The bootstrap was set to 1000 and was run on two threads for the construction of the tree. The protein sequences of OXA-1, OXA-2, OXA-9, OXA-48, OXA-181, OXA-204, OXA-232, and OXA-244 were downloaded as a batch from the NCBI Batch Entrez. The phylogenetic tree was constructed (Figure 19).
PLSDB
IncfIB type plasmid was observed to be predominant in OXA-1, and IncFII was found in GCF_008065415.1 (NCBI accession ID: NZ_CP041928.1). Among CMY-2, IncC was found to be predominant, with GCF_016903775.1 (NCBI accession ID: NZ_CP069963.1) showing IncI2. IncFIB was found in CTX-M-27 and GCF_019334625.1 (NCBI accession ID: NZ_CP079764.1), showing IncFA type. IncC, IncN, and IncFIBtyoe plasmids were found among the IMP-4 genes. IncI, IncFIB, and IncFII were observed in KPC-3. IncHI1B was predominantly found in NDM-1, and GCF_000764615.1 (NCBI accession ID: NZ_CP008933.1) had IncFIB type. IncFIA, IncR, and IncHI1B were found among SHV-11 genes. In VIM-1, IncC was predominant, and GCF_013694425.1 (NCBI accession ID: NZ_CP059314.1) showed type IncR. IncFIB was found to be the most predominant type, occurring in five genes out of the eight understudies, i.e., OXA-1, CTX-M-21, IMP-4, KPC-3, and NDM-1 (Table 5).
MLST
Sequence Types of the carbapenem-resistance genes were identified. Predominantly occurring S.T.s among the strains were observed and reported. Four ST-231 and three ST-11 strains were found to harbor the blaCMY-2 gene. Four genomes containing blaCTX-M-27 belonged to ST-11 and three to ST-48. Four blaIMP-4 harboring strains were ST-340. Twenty-two genomes were found to be ST-258 that had KPC-3 genes. Two genomes belonged to ST-101 that had blaVIM-1. Eleven strains with blaNDM-1 belonged to ST-11, ten to ST-147, and nine to ST-14. Thirty-five genomes with blaOXA-1 were ST-11, and thirty-four were ST-14. One hundred and fifty-six genomes were ST-11 and harbored the blaSHV-11 gene (Figure 20).
Discussions
For studying the co-resistance patterns of the genes, those genes (conferring resistance to other drug classes) that were expressed ≥60% along with the carbapenem-resistant gene were analyzed. Such probability of occurrence suggests that a particular strain could be resistant to more than one class of antibiotics. Eighty-one percent of the genomes expressed fosA3 (fosfomycin resistance gene) in KPC-producing strains. This was the highest recorded co-occurrence with the KPC gene class. A similar pattern was observed in a study that was conducted in a Chinese hospital with a 60.8% co-resistance rate31. Ninety-eight percent of the genomes that produced NDM-1 carbapenemase also showed resistance to glycopeptide antibiotics. OXA-1 producing bacteria expressed resistance to a wide range of antibiotics along with carbapenems. A hundred percent co-resistant rate was found with cephalosporin- resistant CTX-M-2 and diaminopyrimidine-resistant dfrA14 and dfrA23. dfrA14 also confers resistance to rifamycin, and 92% of the genomes expressed this gene. A macrolide-resistant gene, ereA2, was observed with a co-resistance rate of 94.4%, and 96.4% co-resistant APH(3')-VI of aminoglycoside was observed. Within the carbapenem-resistance genes, OXA-1 was found to be co-occurring with NDM class genes with a co-resistance rate of >83%. This is correlated with a study conducted in Dubai, UAE, showing a similar pattern of resistance32. This study is important, especially for clinicians, because it would help them dispense antibiotics to which the bacteria are susceptible. SHV-producing genes were found to co-occur with most of the other carbapenem-resistant genes and other drug class resistant genes and this can be attributed to their ESBL characteristic.
Certain neighboring genes to the resistance-conferring genes could play a role in the mechanism of resistance. To understand how resistance genes function in inactivating an antibiotic, studying the upstream and the downstream genes is indispensable. By studying the genetic context of CMY-2 and CTX-M-27, IS1380/ISEcp1 element was found to occur immediately upstream of CMY-2 and CTX-M-27. Previous studies suggest that ISEcp1 helps in the mobilization of the CMY-2 and CTX-M-27 genes, functioning as a transposon through plasmids in Citrobacter freundii33. ISEcp1 was also found to be important clinically in the spread of ESBLs. ISKra4 has a similar function, and it was found to be occurring immediately downstream of the CTX-M-27 gene. The neighboring genes of IMP-4 included qacG2, aac(61)-Ib4, and catB3. These genes were found downstream of the IMP-4 gene. The blaIMP-4-aacA4-catB3 cassette was found in clinical isolates of Acinetobacter baumanii in Sydney and Melbourne, Australia, suggesting the movement of this cassette through plasmids mediated by homologous recombination34. qacG2 codes for an ammonia compound efflux Small Multidrug Resistance (SMR) transporter. In VIM-1 and OXA-1 genes, intl1-class 1 integron integrase was found in the upstream and the downstream regions, respectively. Integron consists of integrase or size-specific recombination and attI site and acts as a strong promoter in the case of VIM-1. The antibiotic resistance gene cassette integrates with the integron, and the transfer is mediated by homologous recombination35. aac(6')-Ib-cr5, a variant of aac(6')-Ib, found upstream to blaOXA-1 codes for aminoglycoside N-acetyl transferase. It acetylated fluoroquinolones and aminoglycosides, which were found to be the major cause of resistance to aminoglycosides36. The presence of bleMBL downstream to the blaNDM-1 gene was observed, and it confers resistance against bleomycin, an anti-tumor glycopeptide37. Two insertion sequence elements were found downstream of blaNDM-1. Upstream of blaKPC-3, istB-an IS21 putative ATP binding protein is expressed only when there is a tandem repeat of IS21. Immediately after that, istA if IS21 is seen. It cleaves the 31 ends of IS21 tandem repeats elements. Both these genes are required for the formation of cointegration, and the plasmids that carry these cointegrates with other replicons are formed at higher frequencies38. The presence of ɑ/β hydrolase was found immediately downstream.
The phylogenetic study showed that the gene blaOXA-232 is more primitive than the rest of the genes. The short branch length between blaOXA-48, blaOXA-204, and blaOXA-244 suggests that they are more closely related, and the expression pattern could be similar. blaOXA-244 is a derivative of blaOXA-48 and is a weak carbapenemase39. The commonly occurring incompatibility groups of plasmids were analyzed. IncFIB was the most commonly occurring plasmid, followed by IncC. IncFIB was found to be a conjugative plasmid and also involved in the dissemination of antibiotic resistance genes40. Sequence Type analysis using MLST showed the predominant occurrence of ST-11, ST-14, and ST-147 among the genomes. ST-11 is disseminated worldwide and is a single locus variant of ST-258, which is a hyper epidemic clone and is responsible for the dispersion of carbapenem resistance41. ST-147 has been characterized as a high-risk CRKP clone, and the resistance genes were found to be located in multiple plasmids42.
Analyses of all the 82 carbapenem resistance genes were not conducted which limited the scope of studies. Individual resistance gene analysis could give more insights into the mechanisms by which bacteria evade antibiotics. The gathering of data was done only using bioinformatics tools, whereas in vitro characterization and identification of bacteria using Vitek or Minimum Inhibitory Concentrations (MIC) for different antibiotics in vitro were not conducted.
Conclusions
Understanding the resistance determinants like porins, carbapenemases, PBPs, and efflux pumps in order to prolong the lives of infected individuals should be one of the most important goals in combating AMR infections. Synthetic carbapenems that are able to tackle bacterial resistance by bypassing the efflux pumps could be designed. Efflux pump inhibitors have been found to restore the antibiotic activity43, and the focus should also be on coming up with effective inhibitors of Metallo β-lactamases and serine β-lactamases.
References
- Yang P, Chen Y, Jiang S, Shen P, Lu X, Xiao Y. association between antibiotic consumption and the rate of carbapenem-resistant Gram-negative bacteria from China based on 153 tertiary hospitals data in 2014. Antimicrob Resist Infect Control. 2018;7(1):137. [CrossRef]
- Band VI, Satola SW, Burd EM, Farley MM, Jacob JT, Weiss DS. Carbapenem- Resistant Klebsiella pneumoniae Exhibiting Clinically Undetected Colistin Heteroresistance Leads to Treatment Failure in a Murine Model of Infection. mBio. 9(2):e02448-17. [CrossRef]
- Ma YX, Wang CY, Li Y.Y., et al. Considerations and Caveats in Combating ESKAPE Pathogens against Nosocomial Infections. Adv Sci Weinh Baden-Wurtt Ger. 2020;7(1):1901872. [CrossRef]
- Andrey DO, Pereira Dantas P, Martins WBS, et al. An Emerging Clone, Klebsiellapneumoniae Carbapenemase 2-Producing K. pneumoniae Sequence Type 16, Associated With High Mortality Rates in a CC258-Endemic Setting. Clin Infect Dis Off Publ Infect Dis Soc Am. 2020;71(7):e141-e150. [CrossRef]
- Cole M, Baddiley J, Abraham EP.' β-Lactams' as β-lactamase inhibitors. Philos Trans R Soc Lond B Biol Sci. 1980;289(1036):207-223. [CrossRef]
- Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: Past, Present, and Future ▿ . Antimicrob Agents Chemother. 2011;55(11):4943- 4960. 4960. [CrossRef]
- Thienamycin, a new A β-lactam antibiotic I. Discovery,taxonomy, isolation and physical properties. Accessed April 7, 2022. https://www.jstage.jst.go.jp/article/antibiotics1968/32/1/32_1_1/_article/-char/ja/.
- Synthesis and in vitro activity of a new carbapenem, RS-533. Accessed April 8, 2022. https://www.jstage.jst.go.jp/article/antibiotics1968/36/8/36_8_1034/_article/- char/ja/.
- Graham DW, Ashton WT, Barash L, et al. Inhibition of the mammalian .beta.- lactamase renal dipeptidase (dehydropeptidase-I) by Z-2-(acylamino)-3- substituted-propenoic acids. J Med Chem. 1987;30(6):1074-1090. [CrossRef]
- Norrby SR, Alestig K, Björnegård B, et al. Urinary Recovery of N-Formimidoyl Thienamycin (MK0787) as Affected by Coadministration of N-Formimidoyl Thienamycin Dehydropeptidase Inhibitors. Antimicrob Agents Chemother. 1983;23(2):300-307. [CrossRef]
- Oliver A, Levin BR, Juan C, Baquero F, Blázquez J. Hypermutation and the Preexistence of Antibiotic-Resistant Pseudomonas aeruginosa Mutants: Implications for Susceptibility Testing and Treatment of Chronic Infections. Antimicrob Agents Chemother. 2004;48(11):4226-4233. [CrossRef]
- Mandell L. Doripenem: A New Carbapenem in the Treatment of Nosocomial Infection. Clin Infect Dis. 2009;49(Supplement_1):S1-S3. [CrossRef]
- Hugonnet JE, Tremblay LW, Boshoff HI, Barry CE, Blanchard JS. Meropenem- Clavulanate Is Effective Against Extensively Drug-Resistant Mycobacterium tuberculosis. Science. 2009;323(5918):1215-1218. [CrossRef]
- Meletis G. Carbapenem resistance: overview of the problem and future perspectives. Ther Adv Infect Dis. 2016;3(1):15-21. [CrossRef]
- Deris ZZ, Yu HH, Davis K, et al. The Combination of Colistin and Doripenem Is Synergistic against Klebsiella pneumoniae at Multiple Inocula and Suppresses Colistin Resistance in an In Vitro Pharmacokinetic/Pharmacodynamic Model. Antimicrob Agents Chemother. 2012;56(10):5103-5112. [CrossRef]
- Martínez-Martínez L. Extended-spectrum β-lactamases and the permeability barrier. Clin Microbiol Infect. 2008;14(s1):82-89. [CrossRef]
- Pitout JDD, Nordmann P, Poirel L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob Agents Chemother. 2015;59(10):5873-5884. [CrossRef]
- Nordmann P, Cuzon G, Naas T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis. 2009;9(4):228-236. [CrossRef]
- Bebrone C. Metallo-β-lactamases (classification, activity, genetic organization, structure, zinc coordination) and their superfamily. Biochem Pharmacol. 2007;74(12):1686-1701. [CrossRef]
- Yong D, Toleman MA, Giske CG, et al. Characterization of a New Metallo-β- Lactamase Gene, blaNDM-1, and a Novel Erythromycin Esterase Gene Carried on a Unique Genetic Structure in Klebsiella pneumoniae Sequence Type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046-5054. [CrossRef]
- Poirel L, Potron A, Nordmann P. OXA-48-like carbapenemases: the phantom menace. J Antimicrob Chemother. 2012;67(7):1597-1606. [CrossRef]
- Tsilipounidaki K, Athanasakopoulou Z, Müller E, et al. Plethora of Resistance Genes in Carbapenem-Resistant Gram-Negative Bacteria in Greece: No End to a Continuous Genetic Evolution. Microorganisms. 2022;10(1):159. [CrossRef]
- van Duin D, Doi Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence. 2017;8(4):460-469. [CrossRef]
- Schwaber MJ, Klarfeld-Lidji S, Navon-Venezia S, Schwartz D, Leavitt A, Carmeli Y. Predictors of Carbapenem-Resistant Klebsiella pneumoniae Acquisition among Hospitalized Adults and Effect of Acquisition on Mortality. Antimicrob Agents Chemother. 2008;52(3):1028-1033. [CrossRef]
- Suay-García B, Pérez-Gracia MT. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiot Basel Switz. 2019;8(3):E122. [CrossRef]
- Alcock BP, Raphenya AR, Lau TTY, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517-D525. [CrossRef]
- Garcia PS, Jauffrit F, Grangeasse C, Brochier-Armanet C. GeneSpy, a user- friendly and flexible genomic context visualizer. Bioinforma Oxf Engl. 2019;35(2):329-331. [CrossRef]
- Tamura K, Stecher G, Kumar S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol. 2021;38(7):3022-3027. [CrossRef]
- Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293-W296. [CrossRef]
- Galata V, Fehlmann T, Backes C, Keller A. PLSDB: a resource of complete bacterial plasmids. Nucleic Acids Res. 2019;47(D1):D195-D202. [CrossRef]
- Jiang Y, Shen P, Wei Z, et al. Dissemination of a clone carrying a fosA3- harbouring plasmid mediates high fosfomycin resistance rate of KPC-producing Klebsiella pneumoniae in China. Int J Antimicrob Agents. 2015;45(1):66-70. [CrossRef]
- Chen YC, Chen WY, Hsu WY, et al. Distribution of β-lactamases and emergence of carbapenemases co-occurring Enterobacterales isolates with high- level antibiotic resistance identified from patients with intra-abdominal infection in the Asia–Pacific region, 2015–2018. J Microbiol Immunol Infect. Published online July 21, 2021. [CrossRef]
- Verdet C, Gautier V, Chachaty E, et al. Genetic Context of Plasmid-Carried blaCMY-2-Like Genes in Enterobacteriaceae. Antimicrob Agents Chemother. 2009;53(9):4002-4006. [CrossRef]
- Espedido BA, Partridge SR, Iredell JR. blaIMP-4 in Different Genetic Contexts in Enterobacteriaceae Isolates from Australia. Antimicrob Agents Chemother. 2008;52(8):2984-2987. [CrossRef]
- Liao J, Chen Y. Removal of intl1 and associated antibiotics resistant genes in water, sewage sludge and livestock manure treatments. Rev Environ Sci Biotechnol. 2018;17(3):471-500. [CrossRef]
- Vetting MW, Park CH, Hegde SS, Jacoby GA, Hooper DC, Blanchard JS. Mechanistic and Structural Analysis of Aminoglycoside N-Acetyltransferase AAC(6′)-Ib and Its Bifunctional, Fluoroquinolone-Active AAC(6′)-Ib-cr Variant,. Biochemistry. 2008;47(37):9825-9835. [CrossRef]
- Khan AU, Maryam L, Zarrilli R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): a threat to public health. BMC Microbiol. 2017;17(1):101. [CrossRef]
- Reimmann C, Haas D. The istA gene of insertion sequence IS21 is essential for cleavage at the inner 3' ends of tandemly repeated IS21 elements in vitro. EMBO J. 1990;9(12):4055-4063.
- Masseron A, Poirel L, Falgenhauer L, et al. Ongoing dissemination of OXA-244 carbapenemase-producing Escherichia coli in Switzerland and their detection. Diagn Microbiol Infect Dis. 2020;97(3):115059. [CrossRef]
- de Oliveira ÉM, Beltrão EMB, Scavuzzi AML, Barros JF, Lopes ACS. High plasmid variability, and the presence of IncFIB, IncQ, IncA/C, IncHI1B, and IncL/M in clinical isolates of Klebsiella pneumoniae with bla KPC and bla NDM from patients at a public hospital in Brazil. Rev Soc Bras Med Trop. 53:e20200397. [CrossRef]
- Zhao J, Liu C, Liu Y, et al. Genomic characteristics of clinically important ST11 Klebsiella pneumoniae strains worldwide. J Glob Antimicrob Resist. 2020;22:519-526. [CrossRef]
- Gondal AJ, Saleem S, Jahan S, Choudhry N, Yasmin N. Novel Carbapenem- Resistant Klebsiella pneumoniae ST147 Coharboring blaNDM-1, blaOXA-48 and Extended-Spectrum β-Lactamases from Pakistan. Infect Drug Resist. 2020;13:2105-2115. [CrossRef]
- Hirakata Y, Kondo A, Hoshino K, et al. Efflux pump inhibitors reduce the invasiveness of Pseudomonas aeruginosa. Int J Antimicrob Agents. 2009;34(4):343-346. [CrossRef]
Figure 4.
Code for running Macros in M.S. Excel for the analysis of co-occurrence of resistance genes.
Figure 4.
Code for running Macros in M.S. Excel for the analysis of co-occurrence of resistance genes.



Figure 5.
Python code for MLST.
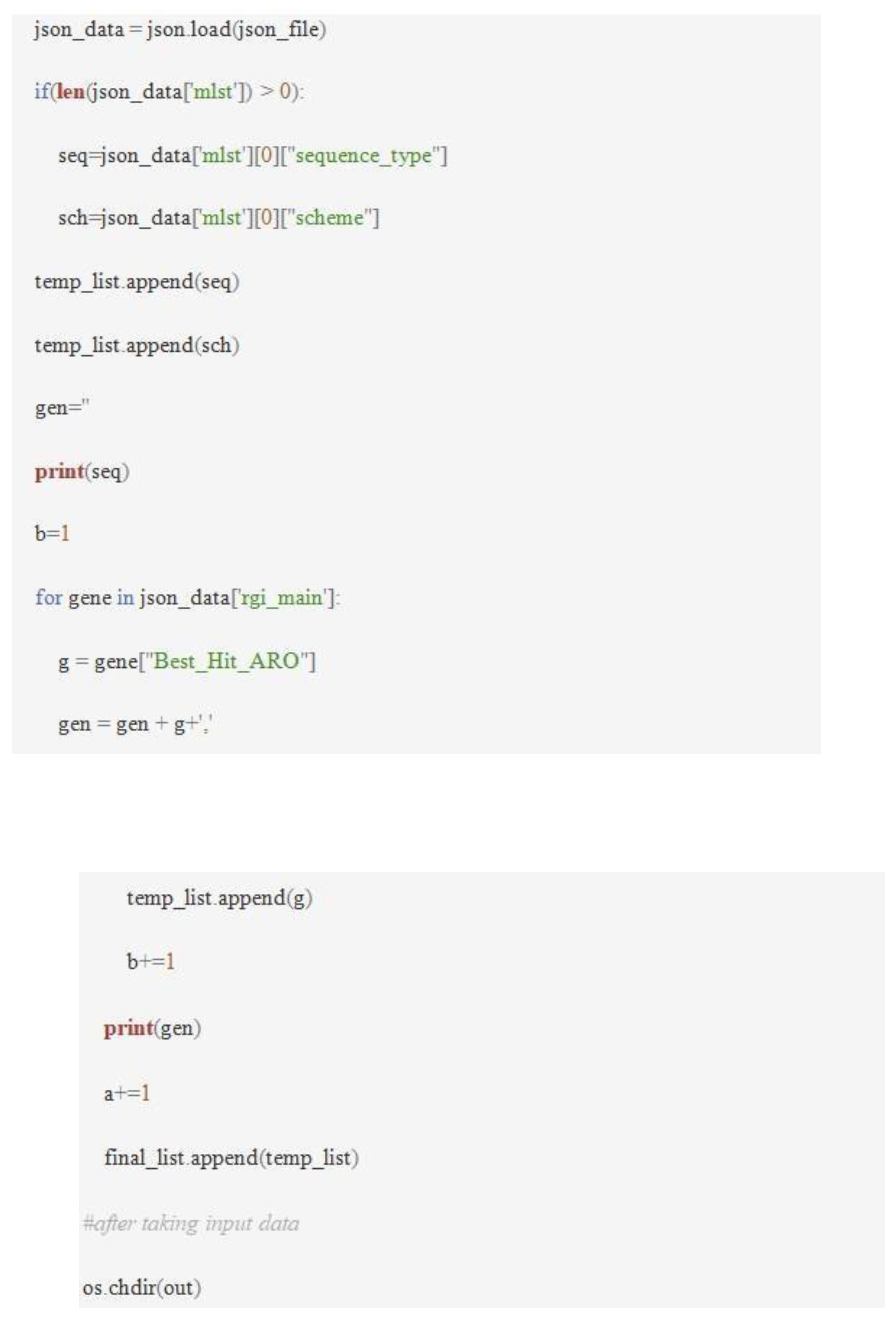
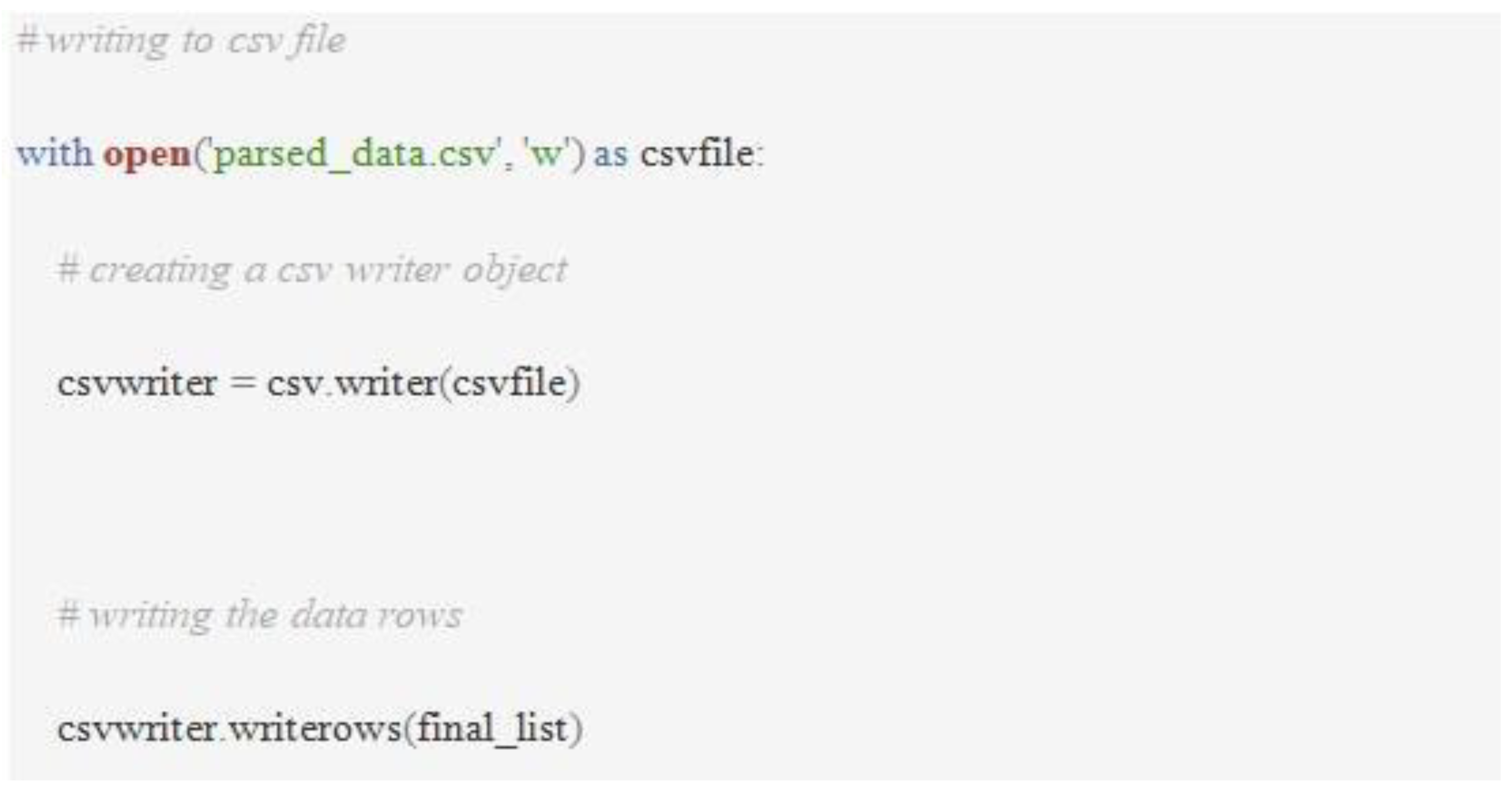
Figure 6.
Distribution of carbapenem resistance genes.
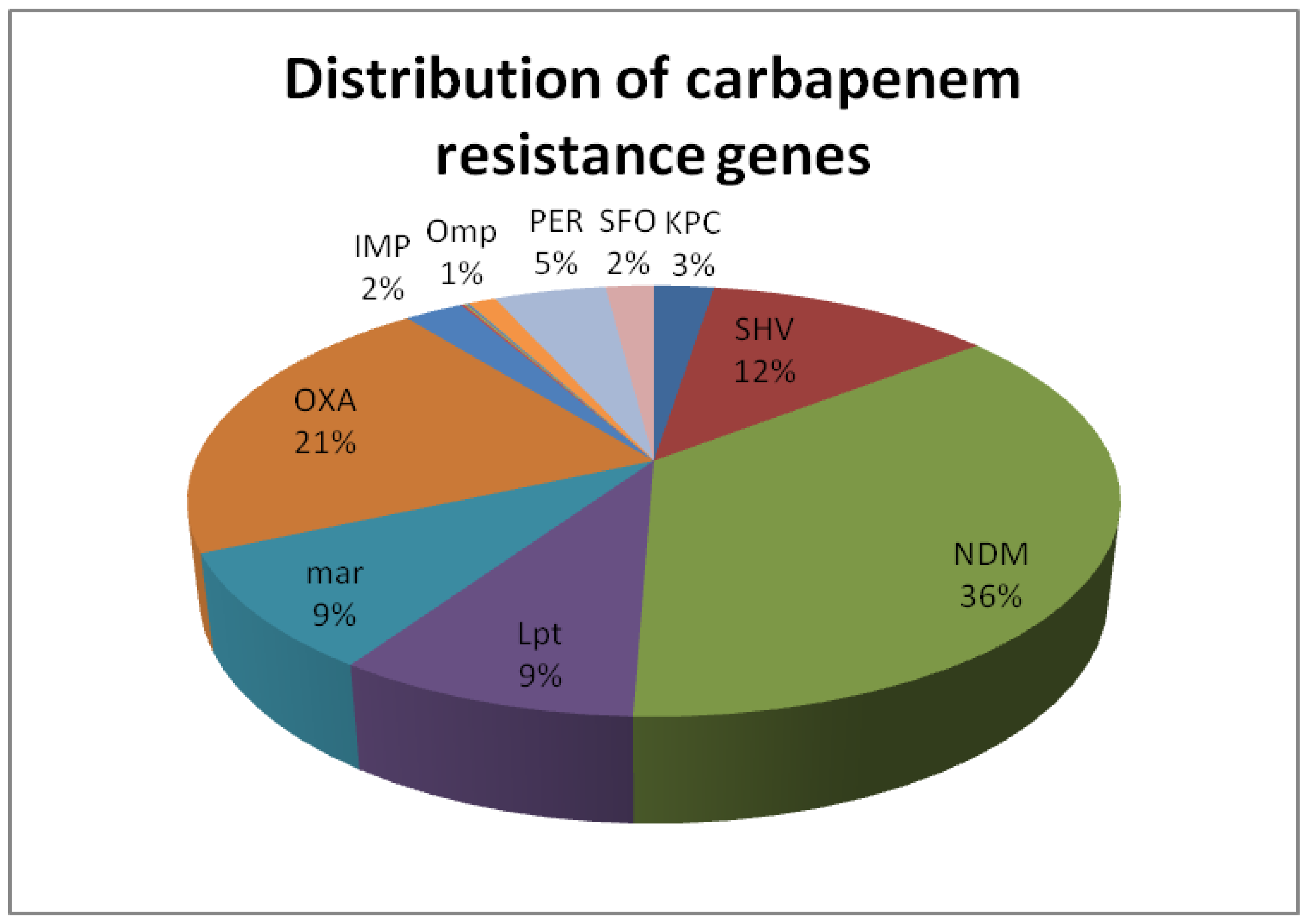
Figure 7.
Co-occurrence of carbapenem-resistance genes with KPC gene class.
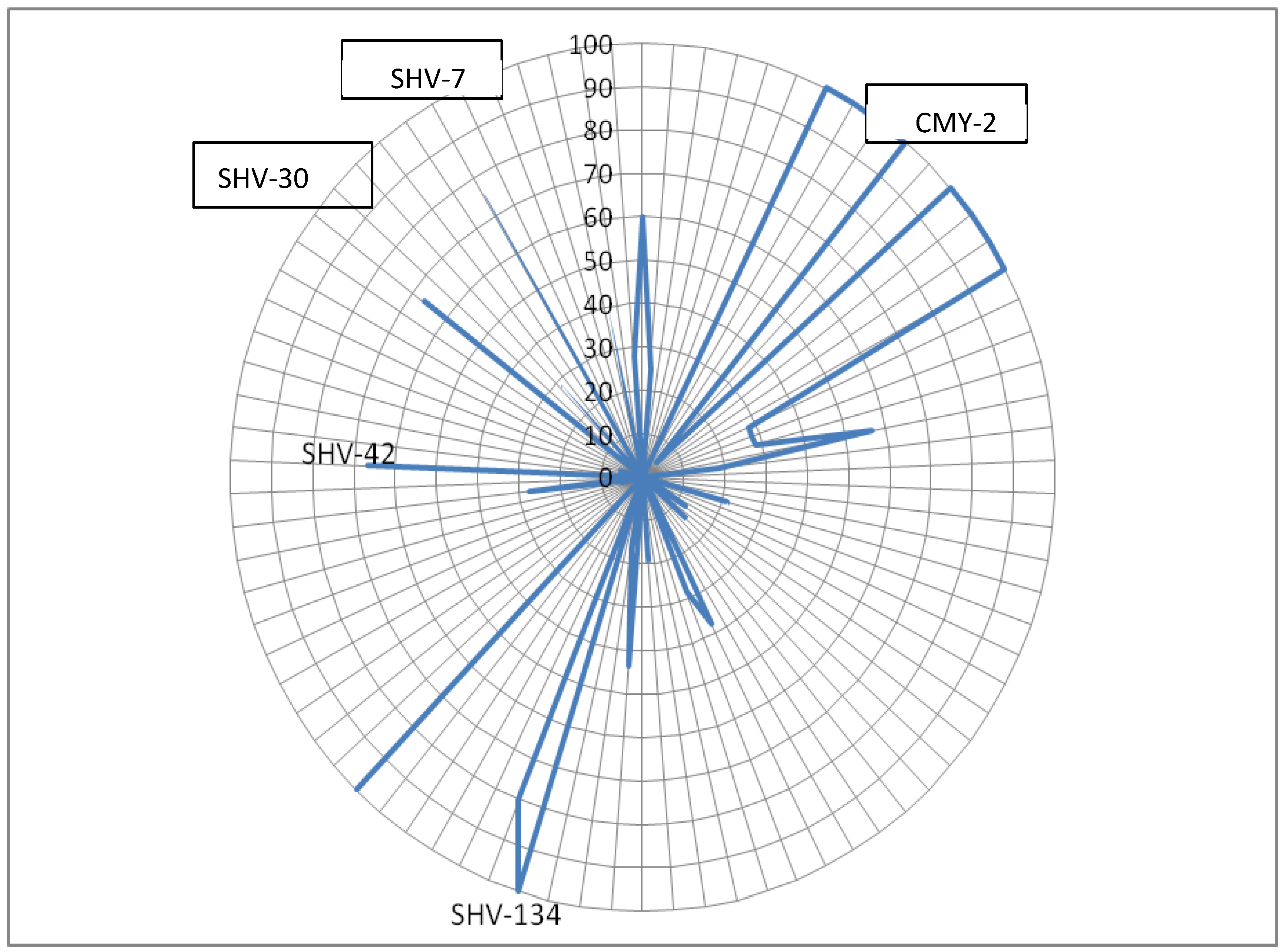
Figure 8.
Co-occurrence of carbapenem-resistance genes with OXA gene class.
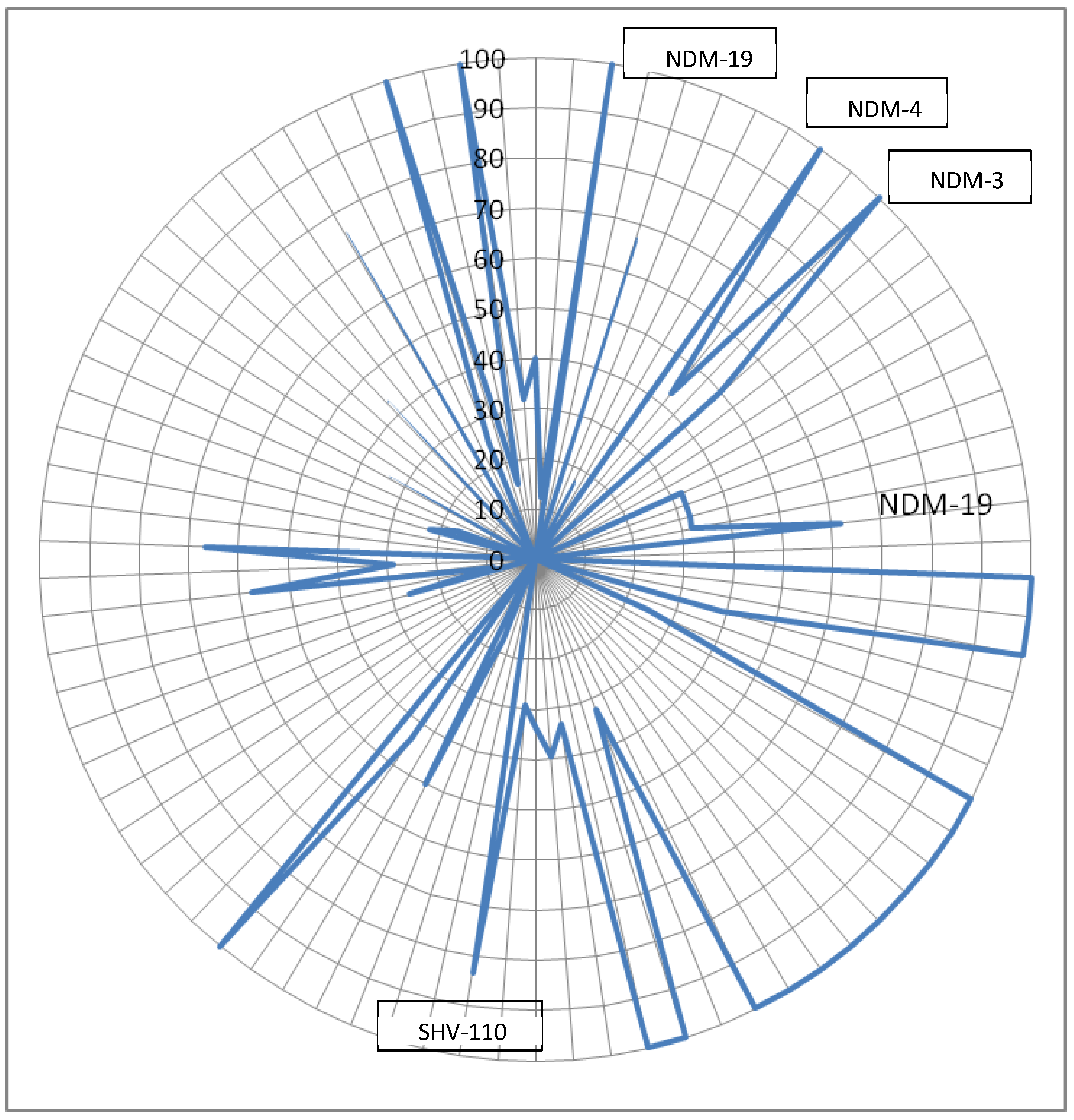
Figure 9.
Genomic context of CMY-2-Klebsiella pneumoniae IR1251 WP_000976514.1 (GCF_017310565.1).
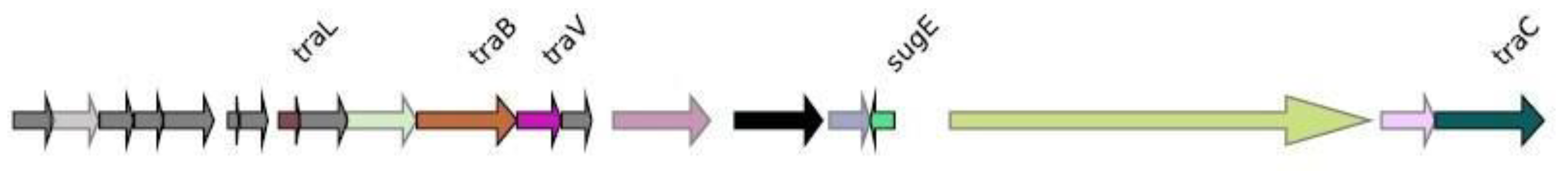
Figure 10.
Genomic context showing tetracycline transporter-Klebsiella pneumoniae JP20 WP_032277257.1 (GCF_019334625.1).
Figure 10.
Genomic context showing tetracycline transporter-Klebsiella pneumoniae JP20 WP_032277257.1 (GCF_019334625.1).

Figure 11.
Genomic context of CTX-M-27-Klebsiella pneumoniae subsp. pneumoniae WP_032277257.1 (GCF_003037615.2).
Figure 11.
Genomic context of CTX-M-27-Klebsiella pneumoniae subsp. pneumoniae WP_032277257.1 (GCF_003037615.2).

Figure 12.
Genomic context of IMP-4-Klebsiella pneumoniae INF157 WP_015060105.1 (GCF_002753075.1).
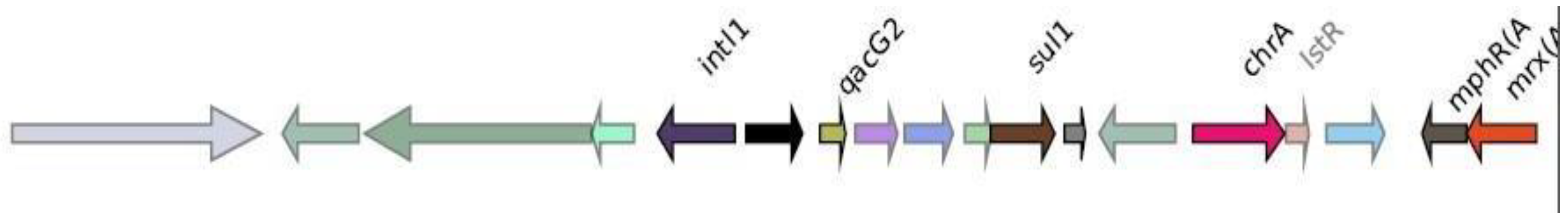
Figure 13.
Genomic context of KPC-3-Klebsiella pneumoniae KPNIH45 WP_004152396.1 (GCF_002918585.2).
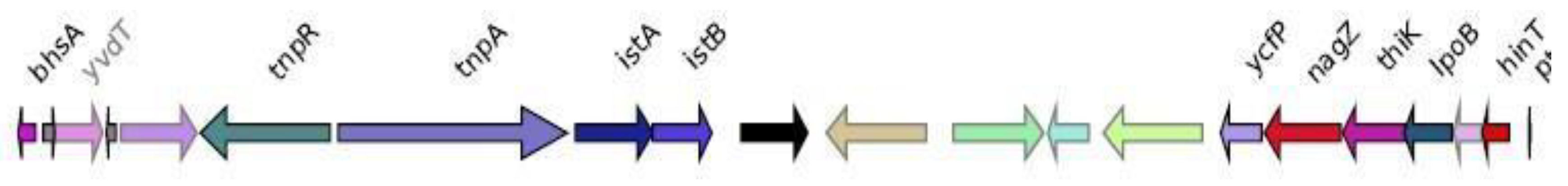
Figure 14.
Genomic context of NDM-1-Klebsiella pneumoniae AR_0068 WP_004201164.1 (GCF_002055855.1).
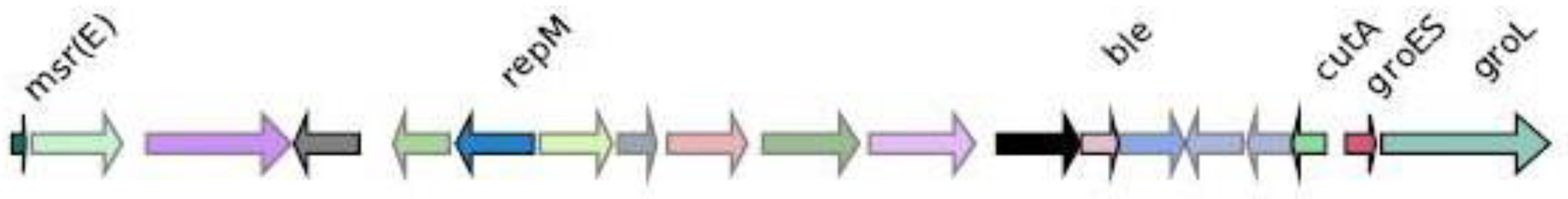
Figure 15.
Genomic context of OXA-1-Klebsiella pneumoniae WP_001334766.1 (GCF_904863225.1).
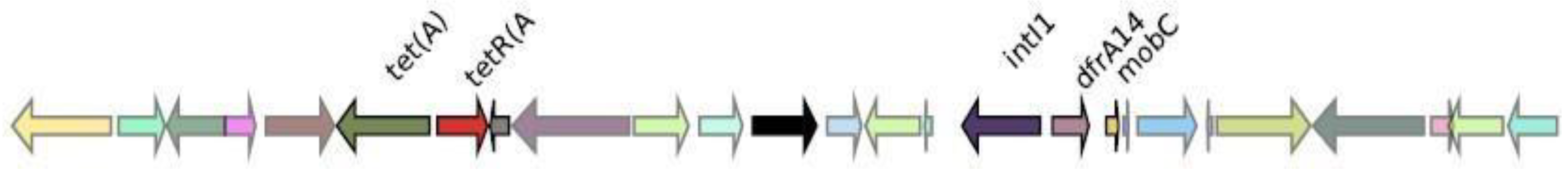
Figure 16.
SHV-11.
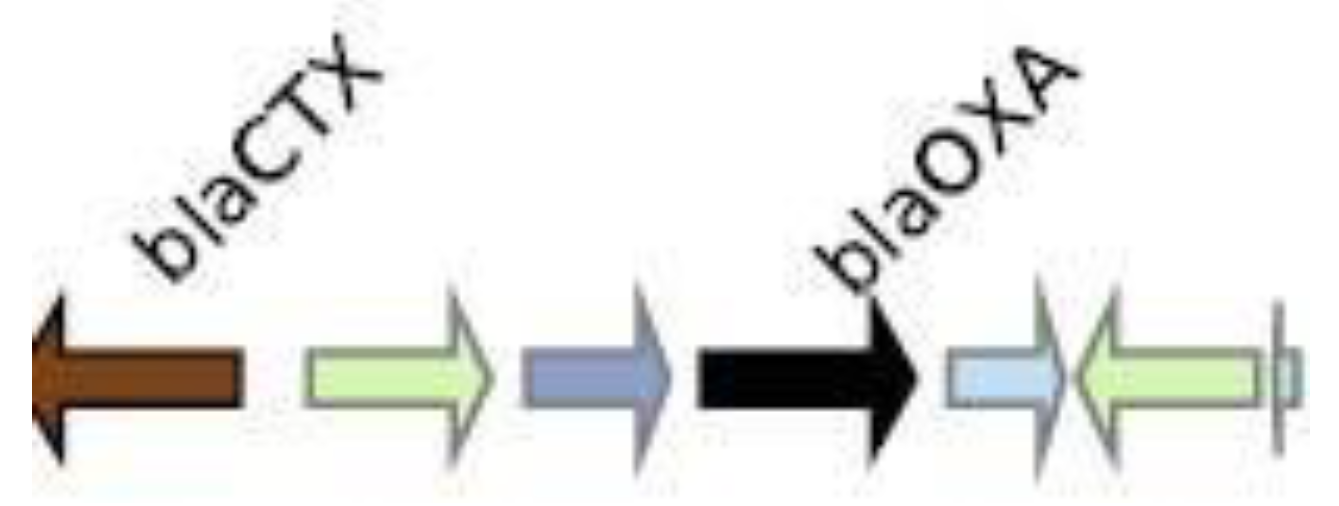
Figure 17.
Genomic context of SHV-11-Klebsiella pneumoniae WP_004176269.1 (GCF_002211665.1).
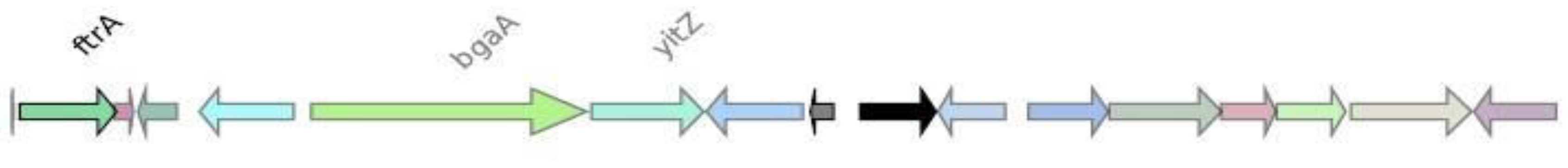
Figure 18.
Genomic context of VIM-1-Klebsiella pneumoniae AR_0135 WP_013263789.1 (GCF_003571545.1).
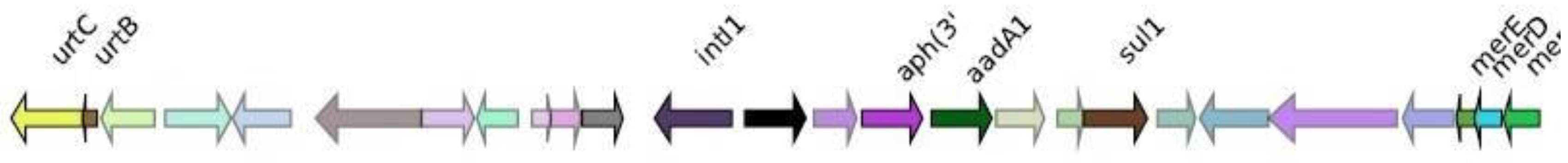
Figure 19.
Phylogenetic tree of the OXA gene class.

Figure 20.
MLST analysis-bar graphs plotted S.T. Vs. Genomes.
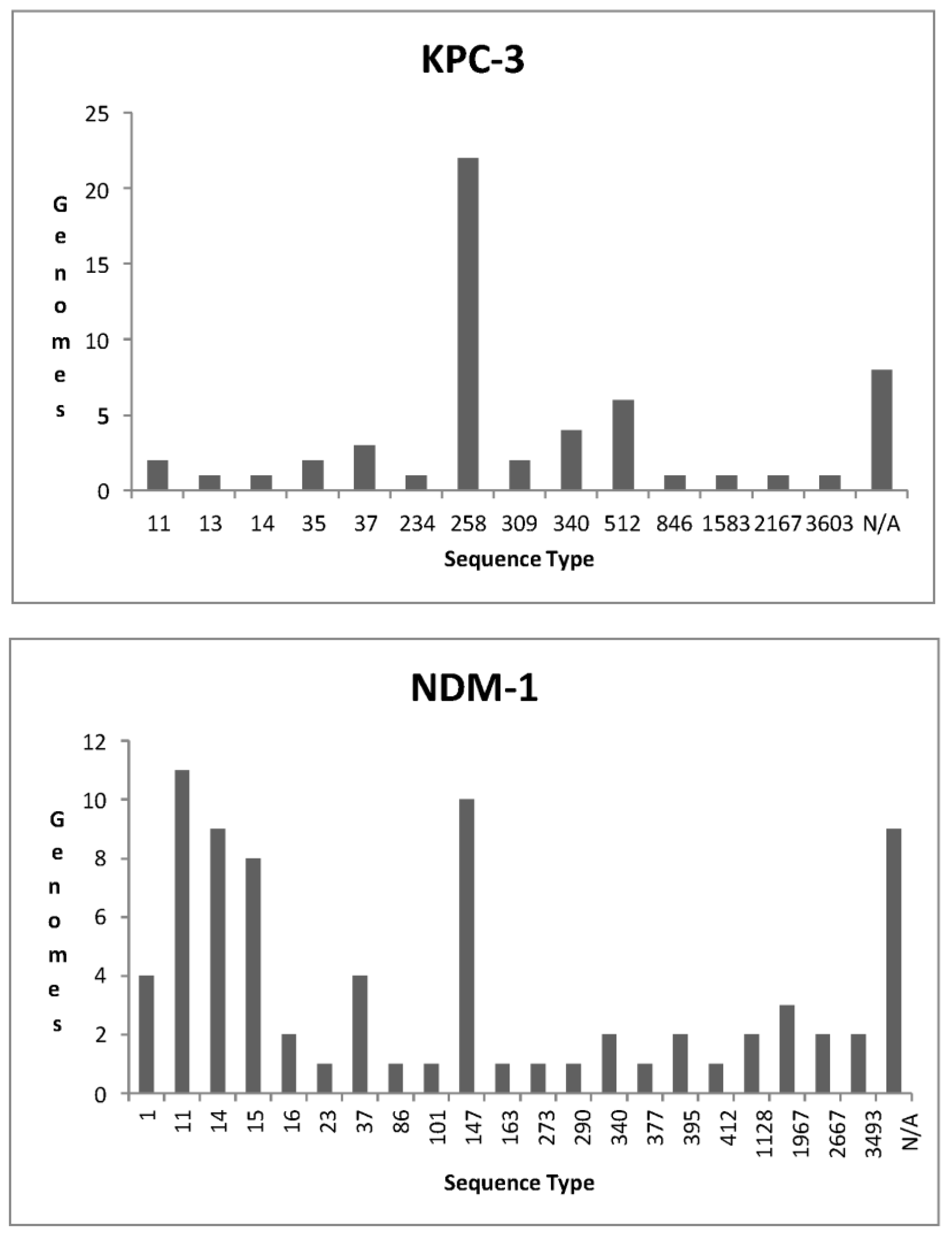
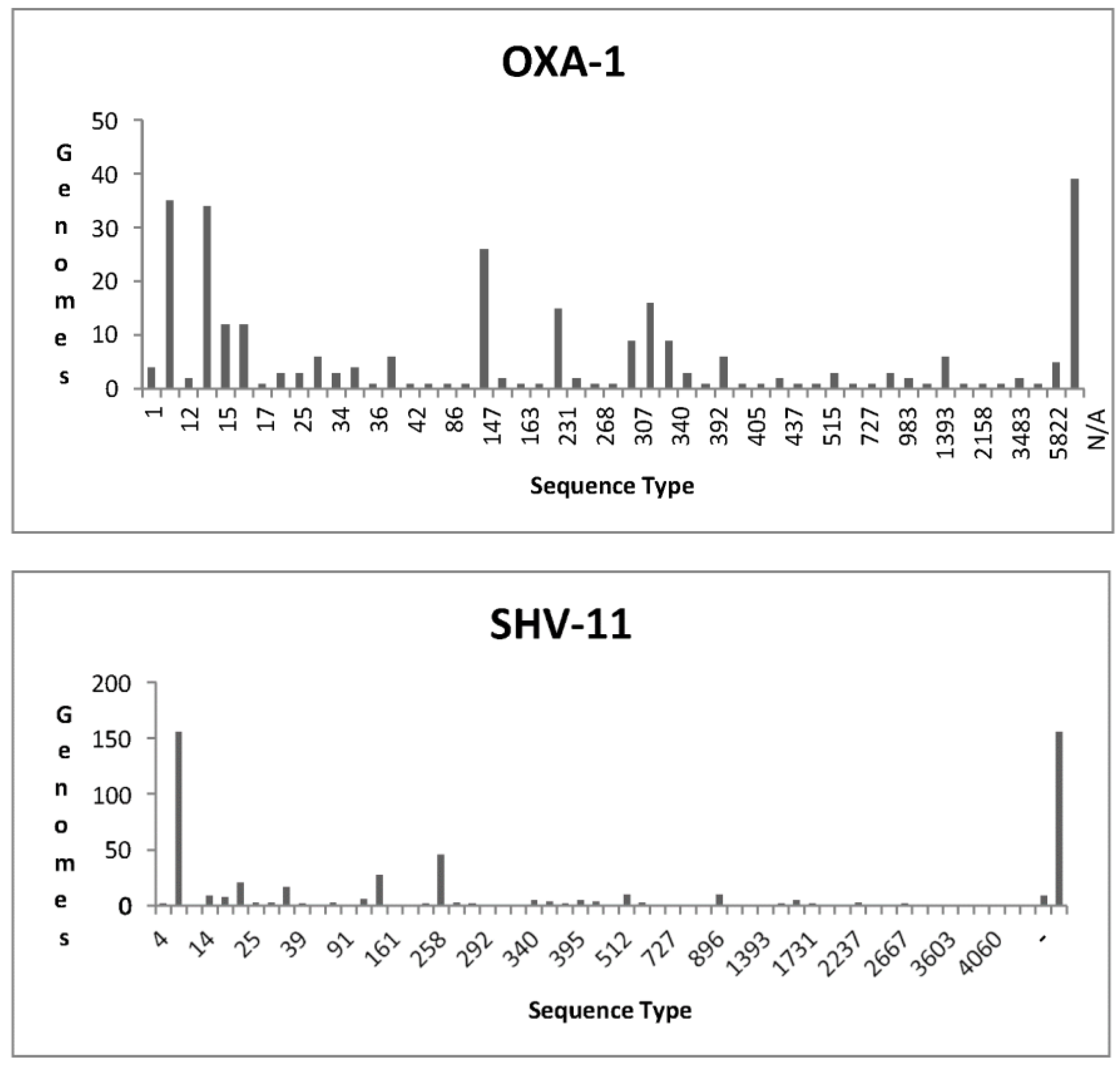

Table 1.
Carbapenemases of different classes.
| KPC | OXA | NDM | IMP | VIM |
|---|---|---|---|---|
| KPC-1 | OXA-1 | NDM-1 | IMP-1 | VIM-1 |
| KPC-3 | OXA-2 | NDM-3 | IMP-4 | VIM-27 |
| KPC-4 | OXA-9 | NDM-4 | IMP-38 | |
| KPC-12 | OXA-10 | NDM-5 | IMP-68 | |
| KPC-14 | OXA-48 | NDM-6 | ||
| KPC-31 | OXA-181 | NDM-7 | ||
| KPC-33 | OXA-204 | NDM-19 | ||
| KPC-40 | OXA-232 | |||
| KPC-74 | OXA-244 |
Table 2.
Co-occurrence of resistance genes with KPC class genes.
| Drug class | Gene/Protein | Percentage |
|---|---|---|
| Aminoglycosides | AAC(3')-IIc | 66.66667 |
| Aminoglycosides | AAC(3')-IV | 61.01695 |
| Aminoglycosides | APH(4')-Ia | 61.01695 |
| Aminoglycosides | LAP-2 | 67.12329 |
| Aminoglycosides | aadA3 | 70.16575 |
| Aminoglycosides | rmtB | 77.41935 |
| Carbapenem | SHV-134 | 80.165 |
| Carbapenem | SHV-30 | 66.66667 |
| Diaminopyrimidine | dfrA25 | 66.66667 |
| Diaminopyrimidine | oqxB | 57 |
| Disinfecting agents and intercalating dyes | qacL | 65.15152 |
| Fluoroquinolones | LAP-2 | 67.12329 |
| Fluoroquinolones | Salmonella serovars gyrB conferring resistance to fluoroquinolone | 79.7619 |
| Fosfomycin | fosA3 | 81.03448 |
| Monobactam | TEM-214 | 66.66667 |
| Penem | SHV-134 | 80.16529 |
| Penem | TEM-214 | 66.66667 |
| Phenicol | catII from Escherichia coli K-12 | 75.92593 |
| Phenicol | cmlA1 | 61.81818 |
| Rifamycin | Escherichia coli rpoB mutants conferring resistance to rifampicin | 66.66667 |
| Rifamycin | LAP-2 | 67.12329 |
| Tetracyclin | LAP-2 | 67.12329 |
| Triclosan | Salmonella enterica gyrA with mutation conferring resistance to triclosan | 72.72727 |
Table 3.
Co-occurrence of resistance genes with NDM class genes.
| Drug class | Genes/Protein | Percentage |
|---|---|---|
| aminoglycoside antibiotic | APH(3')-VI | 92.85714286 |
| diaminopyrimidine antibiotic | dfrA5 | 77.77777778 |
| glycopeptide antibiotic | determinant of bleomycin resistance | 98.11320755 |
Table 4.
Co-occurrence of resistance genes with OXA class genes.
| Drug class | Genes/Proteins | Percentage |
|---|---|---|
| aminoglycoside antibiotic | AAC(3')-IIe | 90.09901 |
| aminoglycoside antibiotic | AAC(6’)-Ib-cr5 | 80.69498 |
| aminoglycoside antibiotic | APH(3')-VI | 96.42857 |
| aminoglycoside antibiotic | APH(3')-VIb | 83.33333 |
| aminoglycoside antibiotic | aadA | 62.26415 |
| aminoglycoside antibiotic | armA | 65.33333 |
| aminoglycoside antibiotic | rmtF | 84.21053 |
| carbapenem | IMP-4 | 66.66667 |
| carbapenem | NDM-1 | 61.84211 |
| carbapenem | SHV-110 | 83.33333 |
| carbapenem | SHV-30 | 66.66667 |
| cephalosporin | CTX-M-194 | 91.66667 |
| cephalosporin | CTX-M-2 | 100 |
| cephalosporin | CTX-M-55 | 66.66667 |
| cephamycin | CMY-4 | 92.30769 |
| diaminopyrimidine antibiotic | dfrA1 | 67.01031 |
| diaminopyrimidine antibiotic | dfrA14 | 61.39535 |
| diaminopyrimidine antibiotic | dfrA23 | 100 |
| diaminopyrimidine antibiotic | dfrA35 | 100 |
| fluoroquinolone antibiotic | AAC(6’)-Ib-cr5 | 80.69498 |
| fluoroquinolone antibiotic | QnrB1 | 80.4878 |
| fluoroquinolone antibiotic | QnrB9 | 66.66667 |
| lincosamide antibiotic | ErmB | 75.67568 |
| macrolide antibiotic | EreA2 | 94.44444 |
| macrolide antibiotic | ErmB | 75.67568 |
| penam | CTX-M-15 | 77.69784 |
| nucleoside antibiotic | SAT-1 | 96.875 |
| rifamycin antibiotic | arr-2 | 92.20779 |
| tetracycline antibiotic | tet(B) | 66.66667 |
| tetracycline antibiotic | tetR | 71.42857 |
Table 5.
Plasmid type distribution and AMR genes carried by them.
| Plasmid type | Gene |
|---|---|
| IncFIA | blaSHV-11 |
| IncFIB | blaOXA-1, blaCTX-M-27, blaIMP-4, blaKPC-3, blaNDM-1 |
| IncFII | blaOXA-1, blaKPC-3 |
| IncHIB | blaNDM-1, blaSHV-11 |
| IncI2 | blaKPC-3 |
| IncFIA | blaSHV-11, blaCTX-M-27 |
| IncC | blaCMY-2, blaIMP-4, blaVIM-1 |
| IncN | blaIMP-4 |
| IncR | blaVIM-1, blaSHV-11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



