
Submitted:
27 January 2026
Posted:
27 January 2026
You are already at the latest version
Abstract
The Brachymystax lenok tsinlingensis is an endangered endemic salmonid species facing severe threats from habitat fragmentation. Effective conservation requires non-invasive monitoring techniques and the identification of key habitat drivers. This study utilized environmental DNA (eDNA) quantitative PCR to estimate the spatial biomass distri-bution of the Brachymystax lenok tsinlingensis in the Zhouzhi Heihe River, integrating surveys of plankton, benthic macroinvertebrates, and multiple physicochemical factors (e.g., water depth, flow velocity, water temperature) to identify critical environmental influences. The results revealed significant spatial heterogeneity in fish biomass, with the highest values observed at river confluences and high-elevation sites. For instance, the confluence of the Heihe River and the Da Manggou River recorded the maximum biomass (1.5 × 10⁻² g/cm²), followed by the Hua Erping site (5.8 × 10⁻³ g/cm²). Among all measured variables, only water depth showed a significant positive correlation with Brachymystax lenok tsinlingensis biomass (r = 0.57). Spatial variations in plankton and benthic community structures reflected a water quality gradient from oligotrophic to eutrophic conditions, but their diversity was not directly linked to fish abundance. These findings indicate that the complexity of in-stream physical habitats—particularly the availability of deep pools—currently plays a more decisive role in the distribution of the Brachymystax lenok tsinlingensis than prey resource diversity. Therefore, conservation efforts should prioritize the restoration of structurally complex river habitats while maintaining high water quality to ensure the long-term survival of this species.
Keywords:
Brachymystax lenok tsinlingensis
; environmental DNA (eDNA)
; biomass distribution
; water depth
; physical habitat complexity
1. Introduction
The Brachymystax lenok tsinlingensis (B. lenok tsinlingensis), commonly referred to as the "flowery fish," is a salmonid species within the genus Brachymystax. Its distribution is primarily confined to the tributary systems of the Wei River on the northern slopes and the Han River on the southern slopes of the Qinling Mountains. This range extends to include tributaries in Zhang Jiachuan, Min Xian, and Gangu counties within the upper Wei River basin of Gansu Province [1]. The evolutionary origin of this species is traced to the Quaternary glacial period approximately 2.5 million years ago. Ancestral salmonids from cold marine environments of the Northern Hemisphere migrated into the Qinling region for spawning. Subsequent abrupt climatic and geological events isolated these fish within the mountain streams. Through a prolonged process of adaptation and speciation, the surviving descendants evolved into the distinct species recognized today as the B. lenok tsinlingensis [2,3]. Intensifying anthropogenic pressures, including habitat encroachment and river fragmentation associated with socio-economic development, have severely impacted this species. As a Chinese endemic and one of the world's southernmost-distributed salmonids—the other being the Sichuan taimen (Hucho bleekeri)—the B. lenok tsinlingensis now faces critical threats. Its populations have become highly fragmented, leading to reduced genetic diversity and a significantly heightened risk of extinction [4,5,6,7]. In recognition of its precarious status, B. lenok tsinlingensis was designated as a Class II National Key Protected Wild Animal in China in 1988. Its conservation priority was reaffirmed with its inclusion in the updated List of National Key Protected Wild Animals (2021 Edition), underscoring the ongoing endangerment of its wild populations [8].
Research on the B. lenok tsinlingensis currently spans multiple scientific domains. Key focuses include morphological analysis [9], ecological studies examining its habitat and life history [5,10], and physiological responses to thermal stress [11,12,13,14]. Applied research has advanced in the fields of artificial propagation, as well as aquaculture techniques [15] and nutritional physiology [16]. From a conservation perspective, work in conservation biology addresses population viability and threat mitigation [17]. Furthermore, molecular biology studies, encompassing genetic diversity, phylogenetics, and adaptive mechanisms, provide a foundational understanding of the species [1,18]. Collectively, these multidisciplinary efforts are essential for developing effective conservation and management strategies for this endangered fish.
In recent years, notable advancements have been achieved in environmental DNA (eDNA) detection technology, with primary applications encompassing population distribution monitoring [19,20], species abundance and biomass estimation [21,22], and life-history tracking [23,24]. Furthermore, studies have shown that eDNA serves as an effective tool for fish biomass assessment not only in controlled aquarium experiments [25,26] but also across diverse aquatic settings, including lentic environments [27,28] and lotic systems [29,30]. To date, however, the application of eDNA to B. lenok tsinlingensis remains limited. Preliminary efforts, including work by our group under aquaculture conditions, have focused on developing and validating biomass assessment methodologies for this species [31].
The Zhouzhi Heihe River, the largest tributary of the Wei River within Zhouzhi County (Xi'an City, Shaanxi Province), spans a broad elevation gradient ranging from 600 to over 3,500 meters. Its basin is predominantly covered by dense forests, with multiple sections designated as national nature reserves. Characterized by ample water supply, high clarity, and minimal pollution, the river provides an ideal habitat for rare fish species such as the B. lenok tsinlingensis [32]. In its lower reaches, the Heihe Reservoir serves as a critical source of drinking water for residents of Xi'an and irrigation water for adjacent agricultural areas [33]. Aquatic community structure and biodiversity are recognized as effective indicators of water quality dynamics [34,35,36]. Pollution-induced disturbances in plankton communities typically involve a decline in sensitive species, an increase in pollution-tolerant taxa, and symptoms of eutrophication [37,38,39,40].
This study utilized environmental DNA quantitative polymerase chain reaction (eDNA-qPCR) to estimate the biomass of the endangered B. lenok tsinlingensis in the Heihe River, Zhouzhi County. Concurrent surveys of plankton and benthic macroinvertebrate community structure were integrated with in situ measurements of key habitat variables, including water temperature, pH, altitude, water depth, turbidity, dissolved oxygen, and flow velocity, enabling the systematic identification of the primary environmental drivers of B. lenok tsinlingensis biomass. Furthermore, the structural characteristics of plankton and benthic assemblages were employed as bioindicators, thereby allowing for a comprehensive assessment of water quality in the Heihe River within Zhouzhi County—a key drinking water source for Xi’an City.
2. Materials and Methods
2.1. Water Sampling Sites
In June 2025, triplicate surface water samples (2 L each) were collected and immediately filtered on-site at fourteen locations along the Heihe River in Zhouzhi County. The locations of the sampling sites are summarized in Table 1, and their spatial distribution across the main channels of the Zhouzhi Heihe River is illustrated in Figure 1. Following filtration, the membrane filters were immersed in 95% ethanol, transported to the laboratory under low-temperature conditions, and promptly stored at −20 °C for subsequent analysis.
2.2. eDNA Extraction and Quantification
Environmental DNA (eDNA) was extracted from the filter using a DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany). Briefly, filters were removed from ethanol, wrapped in analytical-grade filter paper, placed on aluminum foil, and air-dried for 6–7 h. Each dried filter was aseptically cut in half with a sterile razor blade. The halves were transferred into separate 2 mL microcentrifuge tubes, and 500 mL of ATL buffer was added to each tube. The filters were then minced in the buffer using sterile scissors. Digestion was performed with 30 µL of Proteinase K at 56 °C for approximately 48 h. Subsequent steps followed the manufacturer’s protocol, with the following modification: after digestion, 500 mL of AL buffer and 500 mL of absolute ethanol were added to each lysate. Extraction negative controls (filter-free) were processed alongside the samples to monitor for contamination. DNA concentration was determined using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
Quantitative real-time PCR (qPCR) was performed using Premix Ex Taq (Probe qPCR; TaKaRa Bio, Dalian, China) on a Real-Time PCR system. To prevent contamination, sample processing (pre-amplification), PCR setup, and post-amplification analysis were conducted in separate, dedicated rooms. Each 25 µL reaction contained: 12.5 µL of Premix Ex Taq, 8.5 µL of ddH₂O, 10 nM TaqMan probe, and 5 nM of each primer (primer sequences were derived from our previous study [31]). The thermal cycling protocol involved an initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 30 s, annealing at 60 °C for 30 s, and extension at 72 °C for 30 s. All samples were run in four to eight technical replicates, and the mean quantification cycle (Cq) value was used for analysis. An extraction negative control and a no-template control were included with each batch and on every qPCR plate, respectively. A sample was regarded as negative under these conditions if no exponential amplification was observed within 40 cycles. Additionally, primer specificity was verified in silico using Primer-BLAST.
2.3. Methodology for Aquatic Organism Surveys
Aquatic organism surveys were conducted to assess fish resources, plankton communities (phytoplankton and zooplankton), benthic macroinvertebrates, and fish habitats. To minimize potential cross-contamination between molecular and traditional ecological methods, all traditional biological sampling (fish, plankton, and benthos) was performed after the completion of environmental DNA (eDNA) water sample collection and filtration at each site.
2.3.1. Fish Sampling and Processing
Fish were sampled using two baited traps (dimensions: 4.8 m in length, 0.45 m in width, 0.33 m in height), positioned at least 5 m apart at each sampling point. Traps were deployed overnight and retrieved the following day. Captured fish were immediately transferred to a container with MS-222 anesthetic (100 mg/L) for approximately 5 minutes. Following sedation, total length, standard length, and body weight were recorded using digital calipers and an electronic balance. All fish were revived in fresh river water and released at the point of capture immediately after measurement. The fish capture data were obtained from conservation monitoring conducted by the Zhouzhi Fisheries Station in the Heihe River.
2.3.2. Plankton Sampling and Laboratory Analysis
2.3.2.1. Phytoplankton
Phytoplankton samples were collected at a depth of 0.5 m using a water sampler, with 1000 mL collected per sample. Each sample was fixed with 15 mL of Lugol's iodine solution, transferred to a labeled plastic bottle, and stored in the dark at room temperature for transport. In the laboratory, samples were settled for more than 24 h and concentrated to 50 mL. For analysis, a 0.1 mL subsample was examined under an Olympus BX43 microscope equipped with a phytoplankton counting chamber for taxonomic identification and cell density enumeration.
2.3.2.2. Zooplankton
Zooplankton were collected by towing a 64-μm mesh plankton net at a depth of 0.5 m, filtering approximately 20 L of water. The concentrated sample was preserved with 2 mL of formaldehyde in a 100 mL plastic bottle and stored in the dark at room temperature. For analysis, a 1 mL subsample was taken from the concentrate and examined under an Olympus BX43 microscope fitted with a counting chamber for taxonomic identification and enumeration.
2.3.3. Benthic Macroinvertebrate Sampling
Benthic macroinvertebrates were collected from shallow, rocky habitats using a Surber sampler (No. 60 mesh, 250 μm; frame size 30 × 30 cm). A total area of 1 m² was surveyed at each site. Samples were initially sieved on-site, transferred to white sorting trays for preliminary examination, preserved in 99 % ethanol, and stored in 50 mL specimen bottles. In the laboratory, organisms were identified to the lowest feasible taxonomic level (typically species or genus) using a combination of a stereomicroscope and an Olympus BX43 compound microscope. Wet biomass was measured using a precision analytical balance (0.0001 g accuracy), and both abundance (ind./m²) and biomass (g/m²) were subsequently calculated.
2.3.4. Taxonomic References
Taxonomic identification was performed using standard morphological keys and authoritative regional monographs. Phytoplankton were identified primarily to the genus level with reference to Systematics, Classification and Ecology of Freshwater Algae in China [41]. Zooplankton (including rotifers, cladocerans, and copepods) were identified to the lowest possible taxonomic level (typically species or genus) based on Freshwater Rotifera of China [42], Fauna Sinica: Freshwater Cladocera [43], Fauna Sinica: Freshwater Copepoda [44], and supplementary guidance from New Techniques in Microorganism Monitoring [45]. Benthic macroinvertebrates were identified using Studies on Chinese Naididae [46] and the comprehensive text Aquatic Entomology [47]. Although some references are classic works, they remain the standard taxonomic authorities for the freshwater fauna of the study region.
2.3.5. Water Quality Measurements
At all 14 sampling sites, water quality parameters, including temperature, pH, turbidity, dissolved oxygen, and flow velocity, were measured in situ using a portable HORIBA U-52 multiparameter water quality meter (Horiba, Ltd., Japan). In addition, water depth was recorded at the thalweg using a Dingfeng leveling staff (Model 0708, 3 m length, China). All measurements were performed in triplicate, with the mean value of each parameter being used for subsequent analysis.
2.4. Data Processing and Analysis Methods
Biomass was estimated by inputting the raw qPCR Ct values into a pre-established calibration curve published in our prior study [31]:
where y denotes biomass and x represents the Ct value.
This equation, derived from controlled laboratory experiments, allows for the direct translation of Ct values into quantitative biomass estimates. All calculations were performed in MATLAB R2018b, and the results were compiled to generate a biomass value for each sampling site.
To quantify biodiversity, the Shannon-Wiener index (H’) [48], Margalef richness index (D) [49], and Pielou evenness index (J) [50] were applied to the community data of plankton and benthic macroinvertebrates. The corresponding formulas are given below
where Pi is the proportion of individuals of the i-th species in the sample, S denotes the total number of species, and N represents the total number of individuals.
After compiling a raw data matrix of all environmental variables and fish biomass in Excel 2013, we performed a correlation analysis between these variables and the biomass of B. lenok tsinlingensis using MATLAB R2018b. Subsequently, significance testing and redundancy analysis (RDA) were conducted with the vegan package in R to identify the key environmental factors governing the species’ distribution.
3. Results
3.1. Biomass of B. lenok tsinlingensis Using qPCR Assay
This study estimated the biomass density of B. lenok tsinlingensis in the Zhouzhi Heihe River using an environmental DNA (eDNA)-based quantitative PCR (qPCR) approach. Ct values obtained from 14 sampling sites were converted to biomass density via a pre-established regression model (y=2×10¹¹·e⁻¹·¹⁰²ˣ; R²= 0.9987) [31]. The results revealed pronounced spatial heterogeneity in estimated biomass density across river sections. The highest value occurred at the HD site (the confluence of the Heihe River and the Da Manggou River, 1.5×10⁻² g/cm²), followed by Hua Erping site (5.8×10⁻³ g/cm²), Yu Dongquan site (4.4×10⁻³ g/cm²), and Chen Jiagou site (4.2×10⁻³ g/cm²). Intermediate levels were observed in the HT site (the confluence of the Heihe River and the Taiping River, 2.1×10⁻³ g/cm²), Hou Zhenzi site (2.0×10⁻³ g/cm²), and Chang Ping site (1.5×10⁻³ g/cm²). Lower values were recorded in Ban Fangzi site (6.0×10⁻⁴ g/cm²), HH site (the confluence of the Heihe River and the Hubao River , 4.4×10⁻⁴ g/cm²), Hu Bao site (2.1×10⁻⁴ g/cm²), Da Mang site (1.9×10⁻⁴ g/cm²), Ba Mugou site (4.3×10⁻⁵ g/cm²), and Dong site (2.3×10⁻⁶ g/cm²). Notably, no B. lenok tsinlingensis was detected in the Wang Jia site (Figure 2).
To ensure the reliability of the quantitative estimates, pre-defined detection limits were applied. Based on prior assay validation, the limit of quantification (LOQ) was 114.4910 copies/µL. Reliable quantification was achieved for all sites except the Wang Jia site (no amplification) and the Dong site, where the concentration was below the LOQ, confirming the validity of the qPCR-derived estimates (Table 2). These results provide clear insight into the spatial distribution and relative abundance of B. lenok tsinlingensis within the Zhouzhi Heihe River.
In a complementary trap survey, B. lenok tsinlingensis was physically captured at only two locations: one individual in the Ban Fangzi site (20.2 cm total length, 54.2 g body weight) and one in the Chang Ping site (5.1 cm total length, 1.3 g body weight).
3.2. Results of the Survey
3.2.1. Phytoplankton Indicators
Based on water samples collected in July 2025 from 14 sampling sites in the Zhouzhi Heihe River Basin, a total of 26 phytoplankton species were identified and assigned to five phyla: Bacillariophyta, Cyanophyta, Chlorophyta, Cryptophyta, and Dinophyta (Table S1). Bacillariophyta was the most species-rich group (16 species; 61.54% of the total), followed by Cyanophyta (4 species; 15.38%), Chlorophyta (3 species; 11.54%), Cryptophyta (2 species; 7.69%), and Dinophyta (1 species; 3.85%).
Community composition exhibited pronounced spatial heterogeneity. Bacillariophyta dominated both cell density and biomass at most sites (e.g., Da Manggou site and Chang Ping site). Notably, only diatoms were detected at Hua Erping site, HT site, Ban Fangzi site, and Dong site, indicating highly simplified community structures. In contrast, other groups showed marked site-specific enrichment, with Cyanophyta becoming the absolutely dominant group in cell density at Hou Zhenzi site (180,000 cells/L) and Chen Jiagou site (75,000 cells/L) (Table 3). Hou Zhenzi site also exhibited exceptionally high cyanobacterial biomass (1001.5 mg/L) and recorded the highest dinoflagellate biomass in the survey (200.0 mg/L) (Table 4). In addition, Chlorophyta and Cryptophyta were detected at Ba Muping site, while Chlorophyta was also present at both the Wang Jia and Yu Dongquan sites.
Diversity indices showed substantial spatial variation among sites. The Shannon-Weiner index (H′), Margalef richness index (D), and Pielou index (J) differed markedly across the basin. Da Manggou site, Hua Erping site, and Yu Dongquan site exhibited relatively high diversity (H′ >1.3) and evenness (J > 0.84), whereas H′ values were extremely low at Hu Bao site and HD site. Species richness varied considerably, with the lowest D at Hu Bao site (D = 0.0996) and the highest at Hou Zhenzi site (D = 0.8220). However, Hou Zhenzi site exhibited relatively low evenness (J = 0.4849), indicating dominance by a few highly abundant species (Figure 3). Across all sites, the mean values of H′, D, and J were 1.0522, 0.4047, and 0.7155, respectively.
Overall, the phytoplankton communities can be classified into several distinct structural types reflecting different aquatic environmental conditions. Sites such as Da Manggou site, Hua Erping site, and Yu Dongquan site, characterized by high diversity and evenness with minimal cyanobacterial occurrence, indicate clean and relatively stable waters. Hou Zhenzi site displayed typical eutrophic features, with extremely high cyanobacterial and dinoflagellate biomass, the highest richness, but low evenness. Communities at Hu Bao site and the HD site were highly simplified, while Chen Jiagou site was mainly characterized by markedly elevated cyanobacterial cell density. The remaining sites exhibited transitional community patterns, characterized by the coexistence of multiple phyla and intermediate levels of diversity. Collectively, the pronounced spatial differentiation of phytoplankton community structure indicates substantial variability in trophic status and environmental quality across the basin, with clean and eutrophic conditions coexisting.
3.2.2. Zooplankton Indicators
The zooplankton composition across the 14 sampling sites in the Heihe River, Zhouzhi, was species-poor, with only 8 species identified from 3 phyla (Table S2). The overall mean density was extremely low (1.08 ind./L). Cladocerans were completely absent at all sites, while rotifers and copepods occurred at very low mean densities of 0.57 ind./L and 0.22 ind./L, respectively. Their distribution was spatially patchy and sporadic, with detectable abundances recorded only at a few locations (e.g., rotifers: 2 ind./L at Hua Erping site and 3 ind./L at Da Manggou site; copepods: 1 ind./L at Hou Zhenzi site). Biomass followed a similar pattern, averaging only 0.0012 mg/L overall. Mean biomass values of rotifers and copepods were 0.0011 mg/L and 0.0009 mg/L, respectively, with elevated values likewise restricted to isolated sites (e.g., rotifer biomass: 0.0024 mg/L at both Hua Erping and Hu Bao sites; copepod biomass: 0.0030 mg/L at Hou Zhenzi site, Ban Fangzi site, and Chen Jiagou site). Across the vast majority of sites, densities and biomass of all zooplankton groups were at or near the detection limit. Overall, this depauperate, low-abundance zooplankton assemblage reflects a markedly simplified aquatic food web.
3.2.3. Macrobenthos Indicators
A total of 26 benthic species were identified across 14 sampling sites in the Heihe River Basin of Zhouzhi County. The macroinvertebrate community was structurally intact and taxonomically diverse, being dominated by pollution-sensitive aquatic insects. Ephemeroptera (mayflies) emerged as the most abundant group, accounting for approximately 63% of total abundance and represented by 10 genera. Pollution-sensitive indicator taxa such as Ephemeroptera and Trichoptera were prevalent, whereas pollution-tolerant groups like Diptera and Coleoptera did not dominate—a pattern further supported by biomass data (Table S3). This community composition suggests well-oxygenated water, low organic pollution, and generally good water quality.
Biodiversity indices revealed pronounced spatial heterogeneity among the sampling sites. The Shannon–Wiener index (H′) varied from 0.93 at Ba Muping site to 2.31 at Chang Ping site, with Ban Fangzi site also exhibiting high diversity (H′ = 2.27). The Margalef richness index (D) was highest at Ban Fangzi site (3.10) and remained high at Hua Erping site (2.91) and Chang Ping site (2.83), whereas a relatively low value occurred at the HH site (1.16) and Yu Dongquan site (1.13). The Pielou evenness index (J) was generally elevated (>0.71) across most sites, approaching the theoretical maximum at HT site (0.96) and Chang Ping site (0.93). In contrast, Ba Muping site exhibited notably low evenness (0.42), implying possible dominance by a limited number of species and potential environmental stress (Figure 4).
Overall, benthic community composition and biodiversity indices suggest generally clean water conditions and a healthy aquatic habitat in the Heihe River.
3.3. Habitat Factors Influencing the Biomass of B. lenok tsinlingensis
By measuring a suite of abiotic factors (including water temperature, pH, altitude, water depth, turbidity, dissolved oxygen, and flow velocity) as well as biotic factors such as phytoplankton and macrobenthos diversity, key habitat conditions were characterized (Table 5). Correlation analysis between these habitat factors and the biomass of B. lenok tsinlingensis revealed that water depth was the only variable positively associated with both biomass and spatial distribution, with a correlation coefficient of 0.57 (Table 6). Furthermore, water depth was the sole factor approaching statistical significance (0.05 < p < 0.1) (Figure 5), whereas all remaining variables showed no significant relationships. This pattern was further supported by redundancy analysis (RDA) ordination (Figure 6), in which the direction and length of the environmental vectors indicated a strong positive association between water depth and fish biomass. In contrast, flow velocity and the macrobenthos diversity index exhibited moderate negative associations with fish biomass.
4. Discussion
This study employed eDNA technology combined with quantitative PCR to systematically assess the biomass of B. lenok tsinlingensis at 14 sampling sites in the Zhouzhi Heihe River. Concurrently, measurements were taken for both abiotic factors (including water temperature, pH, water depth, turbidity, dissolved oxygen, and flow velocity) and biotic factors (such as plankton and benthic organisms) to characterize habitat conditions. The results revealed significant spatial variation in the biomass of B. lenok tsinlingensis among the sampling sites: the highest biomass was recorded at the HD site, followed by Hua Erping site, Yu Dongquan site, and Chen Jiagou site; intermediate levels were observed at midstream sites, including HT, Hou Zhenzi, and Chang Ping, while relatively low biomass was found at downstream sites such as Ban Fangzi, HH, and Hu Bao. No eDNA signal of B. lenok tsinlingensis was detected at Wang Jia site.
Correlation analysis showed that the biomass of B. lenok tsinlingensis was significantly and positively correlated with water depth (r = 0.57), whereas no significant relationships were observed for the other habitat variables. This finding aligns with previous studies highlighting shaded and dark substrates as preferred habitats for the species, suggesting that light availability may indirectly influence its spatial distribution through its association with water depth [51,52]. Notably, although earlier research suggested that flow velocity, water temperature, elevation, and plankton communities are associated with the distribution of this species [5,53], no significant correlations were found between these factors and biomass in the present study.
The community structure and diversity of plankton and benthic organisms, as direct indicators of aquatic ecosystem health, indirectly influence the biomass of top consumers such as the B. lenok tsinlingensis [34,35,54]. However, in the Zhouzhi Heihe River studied here, no direct or statistically significant correlation was found between the diversity of plankton or benthic organisms and the B. lenok tsinlingensis 's biomass. Plankton communities are effective tools for water quality diagnosis [40,55], and their spatial heterogeneity reveals an environmental gradient from oligotrophic to eutrophic conditions, indirectly affecting the suitable habitat range for this fish. Nonetheless, they are not the direct drivers of its biomass distribution. The benthic macroinvertebrate community overall indicates good water quality and a healthy ecosystem foundation [36], providing an essential prey base. However, spatial variation in its diversity did not directly translate into differences in fish biomass, suggesting that prey availability may not be a limiting resource under current conditions. This implies that biomass patterns likely arise from complex, nonlinear interactions among multiple environmental factors in natural habitats, which cannot be adequately explained by simple linear relationships [10].
Research suggests that river confluences with intense and rapid lateral mixing often show elevated eDNA concentrations [56,57,58,59]. Accordingly, the higher eDNA concentrations observed at the HD site and Chen Jiagou site—attributable to turbulent flow conditions and enhanced hydrodynamic mixing—contrast with the lower biomass recorded at confluences affected by anthropogenic disturbances or subdued hydraulics, such as HH site and HT site. This highlights the role of hydrodynamic processes in regulating the spatial heterogeneity of eDNA.
The Hua Erping site combines suitable water depth with the highest elevation in the basin (1340 m). Previous studies suggest that B. lenok tsinlingensis is typically associated with cold, high-elevation reaches [7,11], which may help explain the relatively high biomass observed at this site. Furthermore, historical survey data confirm that the Hua Erping area is a dominant distribution zone for B. lenok tsinlingensis [60].
In this study, the higher eDNA concentration at Yu Dongquan site, a documented breeding area, was consistent with expectations and aligned with prior survey results [31]. The overall lower biomass at some sites may be primarily attributed to fish dispersal following a flood event in the basin during the previous summer, a hypothesis supported by evidence that storm-induced disturbances can reduce local population abundance [15].
Based on the above, future conservation efforts for the B. lenok tsinlingensis in the Zhouzhi Heihe River should prioritize the protection and restoration of in-stream physical habitat complexity. This entails ensuring the availability of key habitat units, such as deep pools and low-flow areas, while sustaining high water quality throughout the watershed. These measures are essential to safeguard the benthic invertebrate prey base and, consequently, to provide multidimensional support for the sustainable development of the B. lenok tsinlingensis population.
5. Conclusions
In the Zhouzhi Heihe River, an eDNA-based assessment revealed pronounced spatial heterogeneity in the distribution of B. lenok tsinlingensis, with water depth identified as the key habitat factor positively associated with its biomass. While phytoplankton communities reflected a trophic gradient from oligotrophic to eutrophic conditions and the benthic macroinvertebrate community indicated good water quality, no direct correlations were found between fish biomass and the diversity of lower trophic levels. These findings collectively suggest that the availability of structurally complex physical habitats—particularly deeper pools—rather than prey diversity, is the primary determinant of its distribution under current conditions. Therefore, conservation efforts should prioritize restoring in-stream habitat complexity, especially deep-pool refugia, while maintaining high water quality to ensure the long-term survival of this threatened salmonid population.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Phytoplankton data sheet; Table S2: Zooplankton data sheet; Table S3: Benthic macroinvertebrate data sheet.
Author Contributions
Conceptualization, H.Z. (Hu Zhao), H.Z. (Han Zhang), K.Z., J.Z., F.K., Q.W. and H.M.; methodology, H.M.; software, H.Z. (Han Zhang), K.Z., F.K., X.A. and Q.W.; investigation, H.Z. (Hu Zhao), H.Z. (Han Zhang), K.Z., X.D., J.Z., F.K., Q.W. and H.M.; data curation, X.A.; writing—original draft preparation, H.M.; writing—review and editing, H.Z. (Hu Zhao), H.Z. (Han Zhang), K.Z., J.Z., F.K., Q.W. and H.M.; visualization, H.Z. (Hu Zhao) and H.M.; funding acquisition, H.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Shaanxi Academy of Science of China [(grant No. 2023k-30)]. Shaanxi Science and Technology Department, Key R&D Program of Shaanxi Province (2024NC-YBXM-119).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, H.X.; Li, Y.; Liu, X.L. Isolation and characterization of eleven novel microsatellite loci of Brachymystax lenok tsinlingensis, a threatened fish endemic to Shaanxi, China. Conserv. Genet. Resour. 2013, 5, 389–391.
- Zhao, Y.; Zhang, C. Threatened fishes of the world: Brachymystax lenok tsinlingensis Li, 1966 (Salmonidae). Environ. Biol. Fish. 2009, 86, 11-12.
- Xing, Y.C.; Lv, B.B.; Ye, E.Q.; Fan, E.Y.; Li, S.Y.; Wang, L.X.; Zhang, C.G.; Zhao, Y.H. Revalidation and redescription of Brachymystax tsinlingensis Li, 1966 (Salmoniformes: Salmonidae) from China. Zootaxa, 2015, 3962, 191–205.
- Ma, L.Y.; Sun, B.; Yuan, Z.H.; Ren, J.A. The distribution and conservation strategy of Qinling Salmon. M. Agr. Sci. Tech. 2012, 13, 283.
- Wu, J.M.; Yang, H.C.; Shao, J.; Du, H.; Wang, C.Y.; Wei, Q.W. Habitat environmental characteristics of Barchumystax lenok tsinlingensis. Acta. Hydrobiol. Sin. 2017, 41, 214–219.
- Xia, J. G., Ma, Y.J.; Fu, C.; Fu, S.J.; Cooke, S.J. Effects of temperature acclimation on the critical thermal limits and swimming performance of Brachymystax Lenok Tsinlingensis: a threatened fish in Qinling mountain region of China. Ecol. Res. 2017, 32, 61–70.
- Xia, J.G.; Peng, M.R.; Huang, Y.; Elvidge, C.K. Acute warming in winter eliminates chemical alarm responses in threatened Qinling lenok Brachymystax lenok tsinlingensis. Sci. Total. Environ. 2020,142807.
- Zhang, X.M.; Zhao, C.L.; Xie, P.; Feng, G.P.; Chen, P.; Wang, L.X.; Xiong, D.M. Genetic differentiation and local adaptation of Chinese lenok (Brachymystax) populations based on whole genome indels. Acta. Hydrobiol. Sin. 2024, 48, 1724-1735.
- Meng, Y.X.; Wang, G.H.; Xiong, D.M.; Liu, H.X.; Zhang, J.L.; Wang, J.L.; Wang, L.X.; Liu, X.L. The validity of subspecies of Brachymystax lenok tsinlingensis Li based on morphological difference analysis. Acta. Hydrobiol. Sin. 2018, 42, 550-560.
- Tao, Y.Z.; Wang, L.Y.; Huang, Q.F.; Deng C.K.; Chen, F.G.; Xia, J.G. Distribution patterns and habitat environmental characteristics of Brachymystax tsinlingensis larval fish. Acta. Hydrobiol. Sin. 2024, 48, 1716-1723.
- Li, P.; Liu, Q.; Li, J.; Wang, F.; Wen, S.; Li, N. Transcriptomic responses to heat stress in gill and liver of endangered Brachymystax lenok tsinlingensis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 38, 100791.
- Ma, F.; Zhao, L.; Ma, R.; Wang, J.; Du, L. FoxO signaling and mitochondria-related apoptosis pathways mediate tsinling lenok trout (Brachymystax lenok tsinlingensis) liver injury under high temperature stress. Int. J. Biol. Macromol. 2023, 251, 126404.
- Xie, X.B.; Wang, Y.B.; Ma, F.; Ma, R.L.; Du, L.Q.; Chen, X. High-temperature-induced differential expression of miRNA mediates liver inflammatory response in Tsinling Lenok Trout (Brachymystax lenok tsinlingensis). Mar. Biotechnol. 2024, 26, 526–538.
- Wang, Z., Ye, H., Liu, P., Lin, S., Wang, Y., Zhou, Q., Jiang, H., and Shao, J. Surviving the heat: The homeostatic regulation mechanism of endangered Brachymystax tsinlingensis. J. Thermal Biol. 2025,127, 104023.
- Ma, L.D. Analysis of the conservation status and resources investigation of the Qinling lenok in the Tianshui Nature Reserve, Gansu. J. Anim. Sci. Vet. Med. 2024, 43, 43-46.
- Ma, F.; Ma, R.L.; Zhao, L. Effects of antimicrobial peptides on antioxidant properties, non-specifc immune response and gut microbes of Tsinling Lenok Trout (Brachymystax lenok tsinlingensis). Biochem. Genet. 2025, 63, 85–103.
- Zhou, Y.B.; Dong, X.H.; Ju, T.; Gan, L.; Wang, Z.L.; Lian, Y.X.; Zhang, P.; Bai, X.F.; Liu, Q.; Zhang, S.H.; et al. Urgent conservation actions are needed for Qinling Lenok Brachymystax lenok tsinlingensis Li, 1966: enlightenment from model simulations. Ecol. Evol. 2025, 15, e71427.
- Wen, S.; Li, P.; Wang, F; Li, J.L.; Liu, H.X.; Li, N. De novo assembly and microsatellite marker development of the transcriptome of the endangered Brachymystax lenok tsinlingensis. Genes Genom. 2020, 42, 727–73.
- Budd, A.M.; Cooper, M.K.; Le Port, A.; Schils, T.; Mills, M.S.; Deinhart, M.E.; Huerlimann, R.; Strugnell, J.M. First detection of critically endangered scalloped hammerhead sharks (Sphyrna lewini) in Guam, Micronesia, in five decades using environmental DNA. Ecol. Indic. 2021, 127, 107649.
- Otsuki, K.; Hamada, M.; Koizumi, N.; Sakamoto, T.; Nakata, K. Quantitative PCR method to detect an extremely endangered bitterling fish (Rhodeus atremius suigensis) using environmental DNA. Landsc. Ecol. Eng. 2023, 19, 79–86.
- Itakura, H.; Wakiya, R.; Sakata, M.K.; Hsu, H.Y.; Chen, S.C.; Yang, C.C.; Huang, Y.C.; Han, Y.S.; Yamamoto, S.; Minamoto, T. Estimations of riverine distribution, abundance, and biomass of anguillid eels in Japan and Taiwan using environmental DNA analysis. Zool. Stud. 2020, 59, e17.
- Dimond, J.L.; Gathright, B.R.; Bouma, J.V.; Carson, H.S.; Sowul, K. Detecting endangered pinto abalone (Haliotis kamtschatkana) using environmental DNA: comparison of ddPCR, qPCR, and conventional diver surveys. Environ. DNA 2022, 4, 1397-1406.
- Bylemans, J.; Furlan, E.M.; Hardy, C.M.; McGuffie, P.; Lintermans, M.; Gleeson, D.M. An environmental DNA-based method for monitoring spawning activity: A case study, using the endangered Macquarie perch (Macquaria australasica). Methods Ecol. Evol. 2017, 8, 646-655.
- Uchii, K.; Wakimura, K.; Kikko, T.; Yonekura, R.; Kawaguchi, R.; Komada, H.; Yamanaka, H.; Kenzaka, T.; Tani, K. Environmental DNA monitoring method of the commercially important and endangered fish Gnathopogon caerulescens. Limnology 2022, 23, 49–56.
- Takahara, T.; Minamoto, T.; Yamanaka, H.; Doi, H.; Kawabata, Z.I. Estimation of fish biomass using environmental DNA. PLoS ONE 2012, 7, e35868.
- Klymus, K.E.; Richter, C.A.; Chapman, D.C.; Paukert, C. Quantification of eDNA shedding rates from invasive bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. Biol. Conserv. 2015, 183, 77–84.
- Klobucar, S.L.; Rodgers, T.W.; Budy, P. At the forefront: Evidence of the applicability of using environmental DNA to quantify the abundance of fish populations in natural lentic waters with additional sampling considerations. Can. J. Fish. Aquat. Sci. 2017, 74, 2030–2034.
- Lacoursière-Roussel, A.; Côté, G.; Leclerc, V.; Bernatchez, L. Quantifying relative fish abundance with eDNA: A promising tool for fisheries management. J. Appl. Ecol. 2016, 53, 148–1157.
- Levi, T.; Allen, J.M.; Bell, D.; Joyce, J.; Russell, J.R.; Tallmon, D.A.; Vulstek, S.C.; Yang, C.; Yu, D.W. Environmental DNA for the enumeration and management of Pacific salmon. Mol. Ecol. Resour. 2019, 19, 597–608.
- Tillotson, M.D.; Kelly, R.P.; Duda, J.J.; Hoy, M.; Kralj, J.; Quinn, T.P. Concentrations of environmental DNA (eDNA) reflect spawning salmon abundance at fine spatial and temporal scales. Biol. Conserv. 2018, 220, 1–11.
- Zhao, H.; Zhang, H.; Zhang, K.Y.; Deng, J.; Fang, C.; Zhang, J.L.; Kong, F.; Jiang, W.; Wang, Q.J.; Ma, H. Y. Application of Environmental DNA for Assessing the Distribution and Biomass of Brachymystax lenok Tsinlingensis in the Zhouzhi Heihe River. Animals 2025, 15, 977.
- Zhao, H.; Wang, Q.J.; Jiang, W.; Deng, J.; Kong, F.; Zhang, H.X. Analysis of variation in water temperature of rare aquatic wildlife nature reserve of Heihe river. Genomics Appl. Biol. 2017, 36, 1505-1510.
- Ma, H.Y.; Zhang, H.; Deng, J.; Zhao, H.; Kong, F.; Jiang, W.; Zhang, H.X.; Dong, X.G.; Wang, Q.J. Detection the eDNA of Batrachuperus taibaiensis from the Zhouzhi Heihe River Using a Nested PCR Method and DNA Barcoding. Animals 2022, 12, 1105.
- Thakur, R.K.; Jindal, R.; Singh, U. B.; Ahluwalia, A.S. Plankton diversity and water quality assessment of three freshwater lakes of Mandi (Himachal Pradesh, India) with special reference to planktonic indicators. Environ. Monit. Assess. 2013, 185, 8355-8373.
- Jakhar, P. Role of phytoplankton and zooplankton as health indicators of aquatic ecosystem: a review. International journal of innovation studies 2013, 2, 489-500.
- Wang, Y.; Liu, J.J.; Feng, Q.; Liu, W.; Liu, X.D.; Wang, Z.J.; Li, B.L.; Kong, D.X. Macrobenthos community structure and water quality evaluation of Heihe River Basin. Chin. Environ. Sci. 2020, 40, 1305-1319.
- Carvalho, L.; Mcdonald, C.; Dehoyos, C.; Mischke, U.; Phillips, G.; Borics, G.; Poikane, S.; Skjelbred, B.; Solheim, A.L.; Van, W.J.; et al. Sustaining recreational quality of European Lakes: minimizing the health risks from algal blooms through phosphorus control. J. Appl. Ecol. 2013, 50, 315-323.
- Fore, L. S.; Grafe, C. Using diatoms to assess the biological condition of large rivers in Idaho (USA). Freshwater Biol. 2002, 47, 2015-2037.
- Aazami, J.; Maghsodlo, H.; Mira, S. S.; Valikhani, H. Health evaluation of riverine ecosystems using aquatic macroinverte brates: a case study of the Mohammad-Abad River, Iran. Int. J. Environ. Sci. Te. 2020, 17, 2637-2644.
- Xu, J.; Liang, T.; Wang, N.Y.; Qin, K.M.; Li, C.H.; Liu, X.S. Characteristics of community structure of zooplankton and bioassessment of water quality in the Datong Lake in Hunan, China. Ecological Science, 2023, 42, 105–113.
- Hu, H.J.; Li, Y.Y.; Wei, Y.X.; Zhu, H.Z.; Chen, J.Y.; Shi, Z.X. China freshwater algae. Shanghai Science and Technology Press: Shanghai, China, 1980.
- Wang J J. Chinese freshwater rotifers. Science Press: Beijing, China, 1961.
- Jiang, X. Z.; Du, N. S. Zoology of China: freshwater cladocera. Beijing: Science Press: Beijing, China, 1979.
- Shen, J.R. Zoology of China (freshwater copepoda). Science Press: Beijing, China, 1979.
- Shen, Y.F.; Zhang, Z.S.; Gong, X.J.; Gu, M.R.; Shi, Z.X.; Wei, Y.X. A New Technique for Microbiological Monitoring. China Construction Industry Press: Beijing, China, 1990.
- Wang, H. Z. Chinese small earthworm research. Higher Education Press: Beijing, China, 2002.
- Tsuda Matsunae. Aquatic entomology. Hokuryukan Co. , Ltd.: Tokyo, Japan, 1962.
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication. University of Illinois Press: Urbana-Champaign, America, 1963.
- Margalef, D.R. Information theory in ecology. Gen. syst. 1958,3, 36-71.
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370-383.
- Zhang, L.; Lan, K.Y.; Song, R.Q.; Xiong, D.M.; Li, R.J.; Li, J.W.; Jiang, H.B.; Shao, J. Selective behavior of juvenile Brachymystax tsinlingensis depends on substrate color, light intensity, and light color. Prog. Fish. Sci. 2023, 44, 179-187.
- Wang, L. Y.; Xia, J. Y.; Deng, C. K.; Huang, Q. F.; Zhao, H. X.; Tao, Y. Z.; Wu, Q.; Xia, J. G.Comparative studies on light color preference of Qinling lenok Brachymystax tsinlingensis and sympatric fish Phoxinus lagowskii in different life history stages.Acta. Ecol. Sin. 2024, 44, 7859-7870..
- Huang, Q.F.; Deng, C.K.; Xia, J.Y.; Yan, H.J.; Xia, J.G. Geometric morphology of Brachymystax tsinlingensis and sympatric Phoxinus lagowskii: life-history stage effects and interspecific differences. Chinese Journal of Ecology, 2024, 43, 922-929.
- Dalia, S.V.; Satheesh, K.P.; Pillai, N.G.K. Biodiversity and seasonal variation of benthic macrofauna in Minicoy Island, Lakshadweep, India. Acta. Oceanol. Sin. 2014, 33, 58-73.
- Reavie, E.D.; Jichab, T.M.; Angradib, T.R.; Bolgrien, D.W.; Hill, B.H. Algal assemblages for large river monitoring: Comparison among biovolume, absolute and relative abundance metrics. Ecol. Indic. 2010, 10, 167-177.
- Laporte, M.; Bougas, B.; Champoux, O.; Morin, J.; Côté, G.; Paradis, Y.; Bernatchez, L. Caged fsh experiment and hydrodynamic bidimensional model highlight the importance to consider advection-difusion dilution in environmental DNA studies in large rivers. Environ. DNA 2020, 2, 362–372.
- Wood, Z.T.; Lacoursière-Roussel, A.; LeBlanc, F.; Trudel, M.; Kinnison, M.T.; Garry M.C.; Pavey, S.A.; Gagné, N. Spatial heterogeneity of edna transport improves stream assessment of threatened salmon presence, abundance, and location. Front. Ecol. Evol. 2021, 650717.
- Van, D.C.; Everts, T.; Neyrinck, S.; Brys, R. Experimental assessment of downstream environmental DNA patterns under variable fish biomass and river discharge rates. Environ. DNA 2023, 5, 102–116.
- Urabe, H., Mizumoto, H., Tsuda-Yamaguchi, F.; Araki, H. Spatial heterogeneity of eDNA concentration as a predictor of small biomass of fish in a mountain stream. Limnology 2025, 26, 223–233.
- Zhang, J.L.; Bian, K.; Jin, T.Z.; Gou, N.N.; Shen, H.B.; Zhang, J.Y.; Wang, K.F.; Ding, Q.Z. Investigation on status of fish resources in Qinling Heihe river basin. Freshw. Fish 2016, 46, 103–108.
Figure 1.
Collection sites for B. lenok tsinlingensis eDNA in the Zhouzhi Heihe River (the map in the top-left corner shows that the red area within China represents the habitat of B. lenok tsinlingensis. This figure was generated using ArcGIS 10.8).
Figure 1.
Collection sites for B. lenok tsinlingensis eDNA in the Zhouzhi Heihe River (the map in the top-left corner shows that the red area within China represents the habitat of B. lenok tsinlingensis. This figure was generated using ArcGIS 10.8).
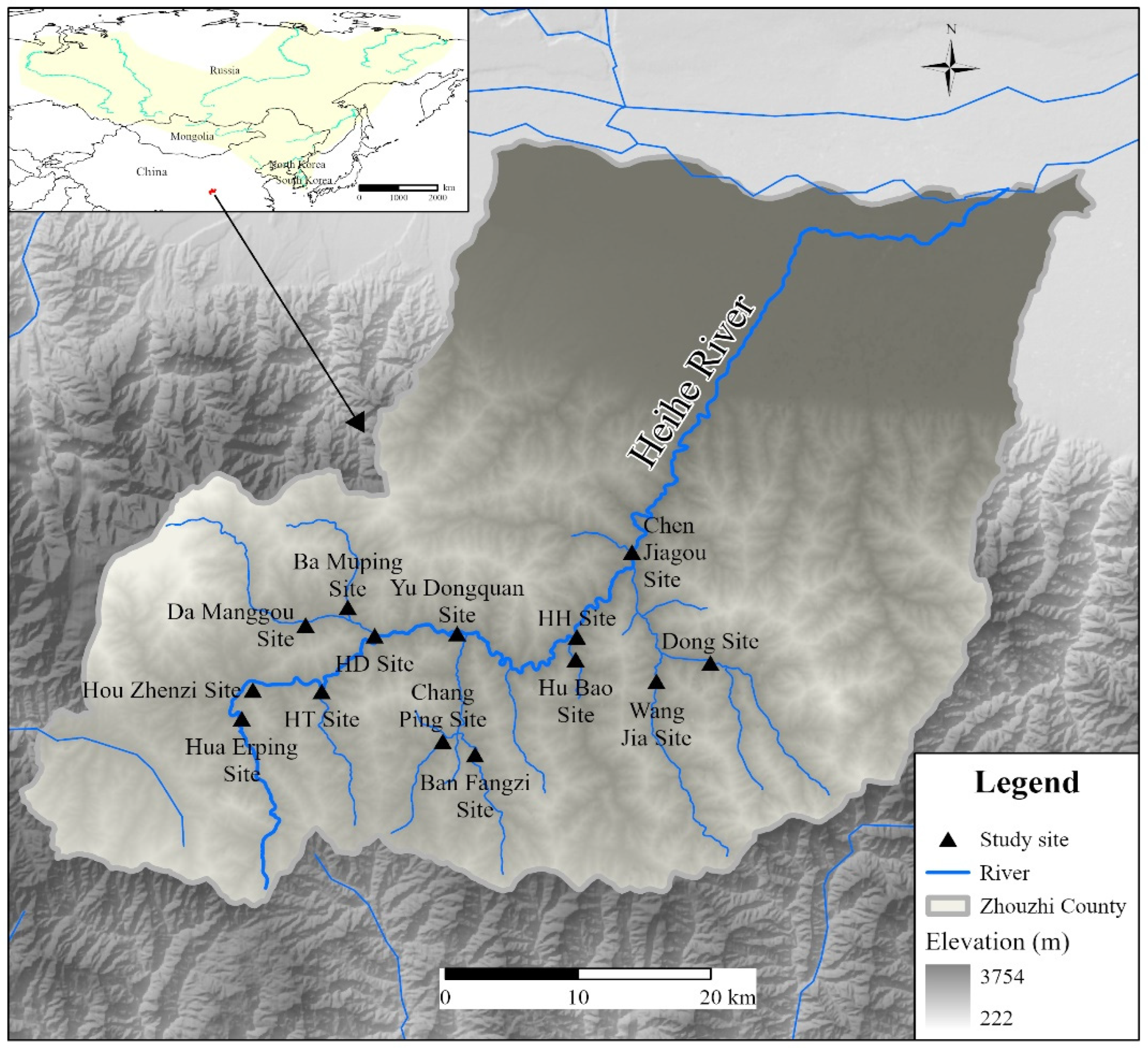
Figure 2.
Biomass of B. lenok tsinlingensis in the Zhouzhi Heihe River.
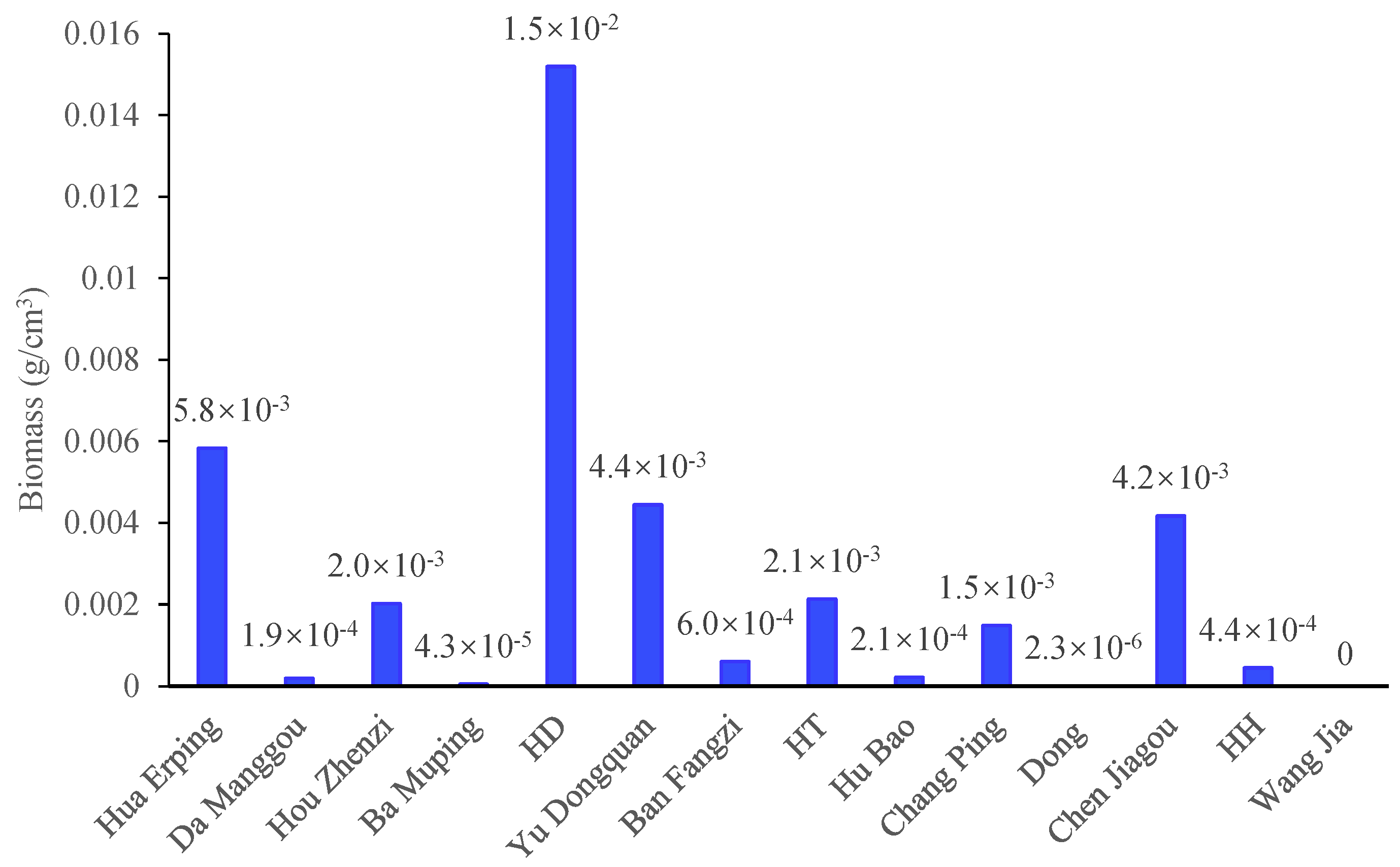
Figure 3.
Phytoplankton diversity indices at different sites in the Zhouzhi Heihe River.
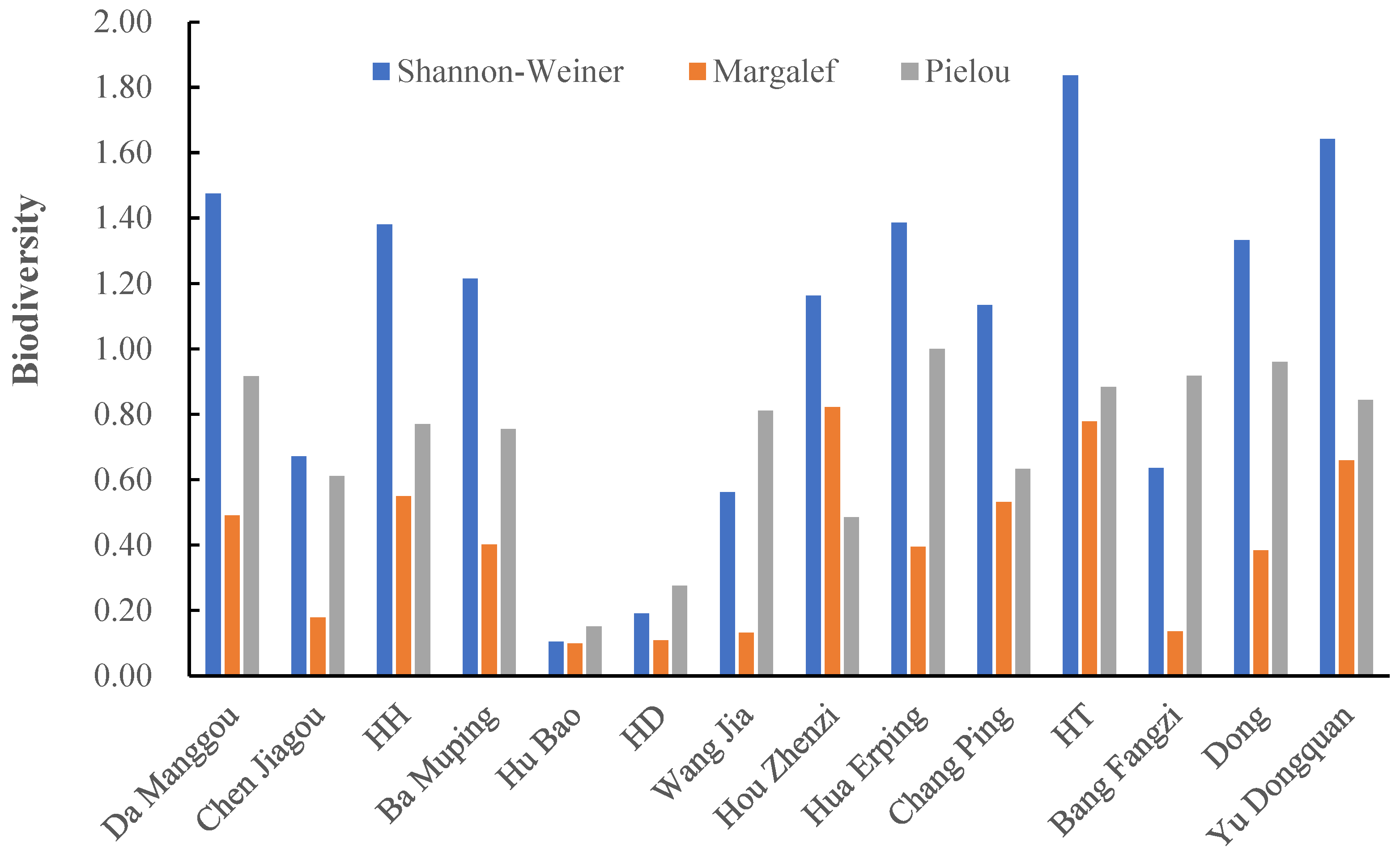
Figure 4.
Macrobenthos biodiversity indicator diagram of the Zhouzhi Heihe River.
Figure 5.
Significance of correlations between B. lenok tsinlingensis biomass and environmental factors. (T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.).
Figure 5.
Significance of correlations between B. lenok tsinlingensis biomass and environmental factors. (T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.).
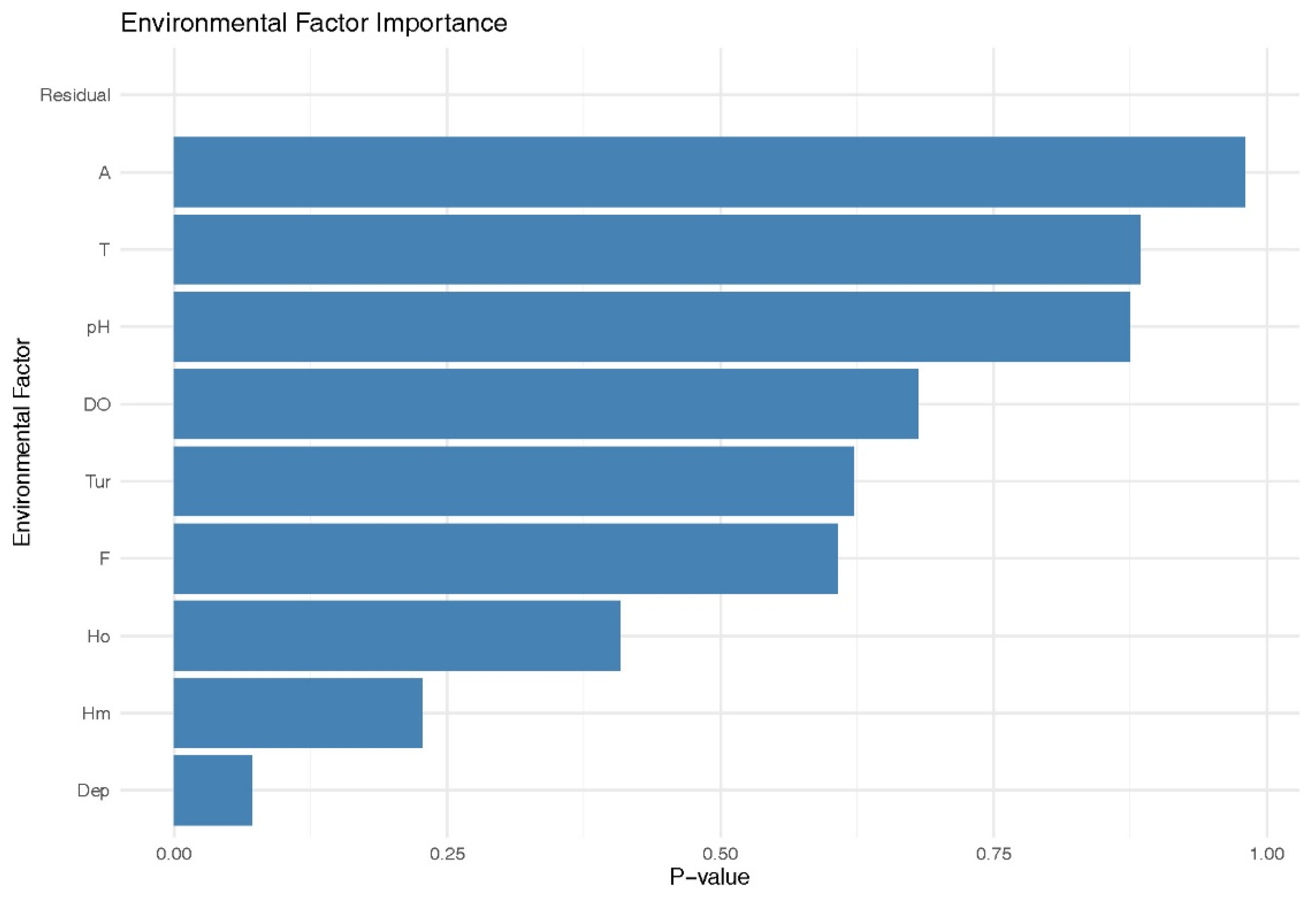
Figure 6.
Redundancy analysis between B. lenok tsinlingensis biomass and physiochemical factors in Heihe River. (T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.).
Figure 6.
Redundancy analysis between B. lenok tsinlingensis biomass and physiochemical factors in Heihe River. (T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.).
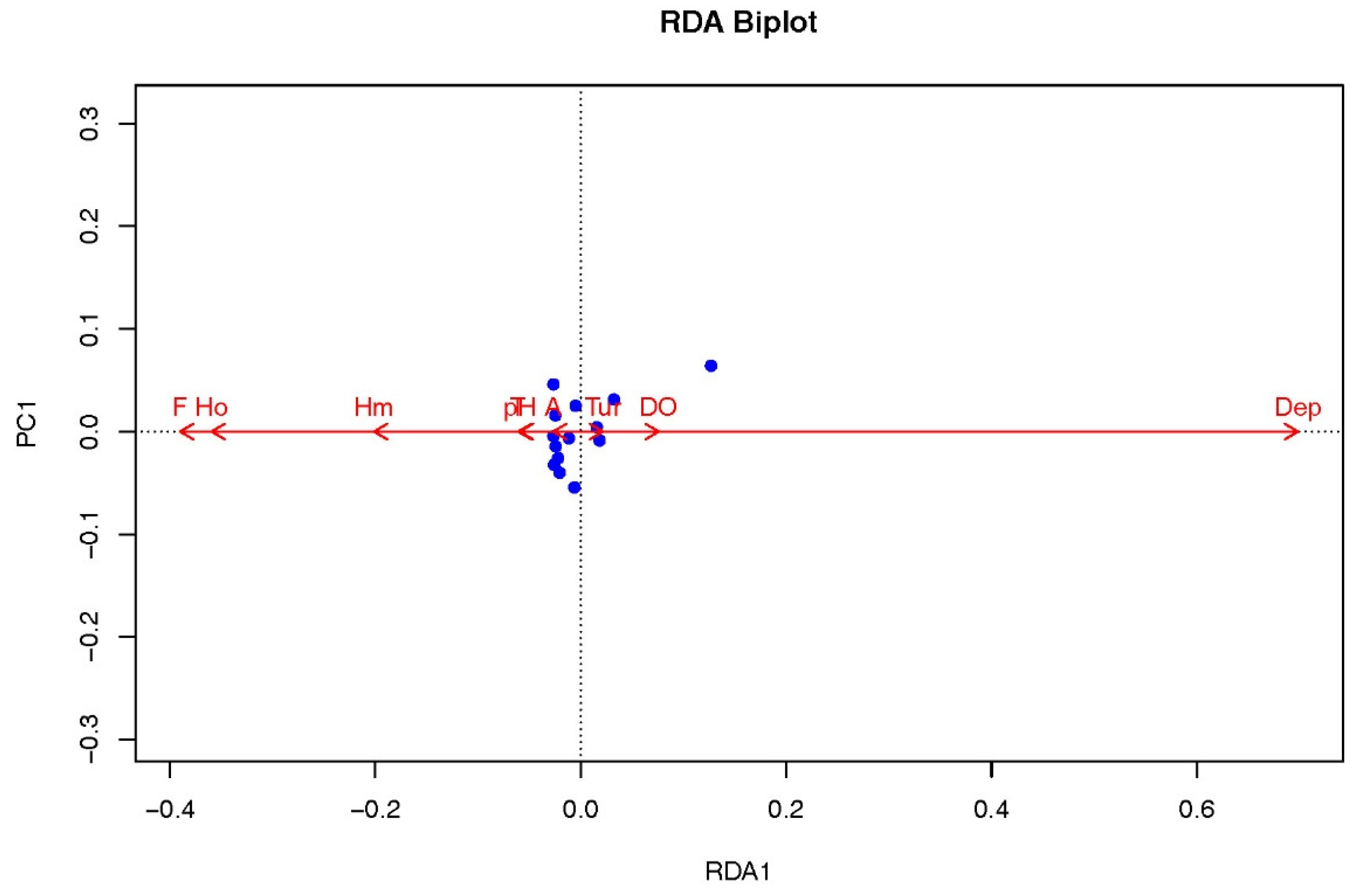

Table 1.
Information regarding the sites where water was sampled for eDNA analysis.
| Number | Location | Coordinates | Altitudes |
| 1 | Hua Erping site | 33.826553 N, 107.832346 E | 1340 m |
| 2 | Hou Zhenzi site | 33.846789 N, 107.840567 E | 1288 m |
| 3 | HT site | 33.847261 N, 107.896643 E | 1170 m |
| 4 | HD site | 33.887228 N, 107.938330 E | 1077 m |
| 5 | Da Manggou site | 33.893200 N, 107.881854 E | 1265 m |
| 6 | Ba Muping site | 33.906418 N, 107.915733 E | 1202 m |
| 7 | Yu Dongquan site | 33.890361 N, 108.005389 E | 963 m |
| 8 | Chang Ping site | 33.814817 N, 107.996285 E | 1158 m |
| 9 | Ban Fangzi site | 33.806271 N, 108.022873 E | 1219 m |
| 10 | HH site | 33.890367 N, 108.102878 E | 819 m |
| 11 | Hu Bao site | 33.874491 N, 108.102443 E | 872 m |
| 12 | Wang Jia site | 33.860821 N, 108.168671 E | 995 m |
| 13 | Dong site | 33.874596 N, 108.212104 E | 981 m |
| 14 | Chen Jiagou site | 33.950493 N, 108.145729 E | 676 m |
Note: HT site, the confluence of the Heihe River and the Taiping River; HD site, the confluence of the Heihe River and the Da Manggou River; HH site, the confluence of the Heihe River and the Hubao River.
Table 2.
Quantification of B. lenok tsinlingensis eDNA using qPCR. The data include the CT value and the average concentrations of B. lenok tsinlingensis eDNA in the Zhouzhi Heihe River.
Table 2.
Quantification of B. lenok tsinlingensis eDNA using qPCR. The data include the CT value and the average concentrations of B. lenok tsinlingensis eDNA in the Zhouzhi Heihe River.
| Number | location | CT Value | eDNA concentration(copy/µL) |
| 1 | Hua Erping site | 28.281 | 5286.044 |
| 2 | Da Manggou site | 31.386 | 788.639 |
| 3 | Hou Zhenzi site | 29.243 | 2932.651 |
| 4 | Ba Muping site | 32.744 | 343.253 |
| 5 | HD site | 27.412 | 9001.277 |
| 6 | Yu Dongquan site | 28.528 | 4543.867 |
| 7 | Ban Fangzi site | 30.349 | 1488.463 |
| 8 | HT site | 29.193 | 3022.873 |
| 9 | Hu Bao site | 31.294 | 834.495 |
| 10 | Chang Ping site | 29.522 | 2471.598 |
| 11 | Dong site | 35.393 | 67.698 |
| 12 | Chen Jiagou site | 28.586 | 4385.650 |
| 13 | HH site | 30.621 | 1260.431 |
| 14 | Wang Jia site | None | None |
Table 3.
Phytoplankton cell density by taxonomic group at selected sites in Zhouzhi Heihe River.
| Cell Density(cells/L) | Cyanobacteria | Chlorophyta | Bacillariophyta | Cryptophyta | Pyrrophyta |
| Da Manggou site | 0 | 0 | 3500 | 0 | 0 |
| Chen Jiagou site | 75000 | 0 | 500 | 0 | 0 |
| HH site | 5000 | 0 | 4000 | 0 | 0 |
| Ba Muping site | 15000 | 5000 | 500 | 500 | 0 |
| Hu Bao site | 22500 | 0 | 500 | 0 | 0 |
| HD site | 10000 | 0 | 500 | 0 | 0 |
| Wang Jia site | 0 | 500 | 1500 | 0 | 0 |
| Hou Zhenzi site | 180000 | 7000 | 4500 | 0 | 500 |
| Hua Erping site | 0 | 0 | 2000 | 0 | 0 |
| Chang Ping site | 0 | 0 | 12000 | 0 | 0 |
| HT site | 0 | 0 | 8000 | 0 | 0 |
| Ban Fangzi site | 0 | 0 | 1500 | 0 | 0 |
| Dong site | 0 | 0 | 2500 | 0 | 0 |
| Yu Dongquan site | 0 | 500 | 8000 | 500 | 0 |
Table 4.
Phytoplankton biomass by taxonomic group at selected sites in Zhouzhi Heihe River.
| Biomass(mg/L) | Cyanobacteria | Chlorophyta | Bacillariophyta | Cryptophyta | Pyrrophyta |
| Da Manggou site | 0 | 0 | 901.4 | 0 | 0 |
| Chen Jiagou site | 99.8 | 0 | 5.0 | 0 | 0 |
| HH site | 16.9 | 0 | 305.6 | 0 | 0 |
| Ba Muping site | 38.9 | 7.5 | 5.0 | 0.5 | |
| Hu Bao site | 76.3 | 0 | 5.0 | 0 | 0 |
| HD site | 33.9 | 0 | 5.0 | 0 | 0 |
| Wang Jia site | 0 | 20.0 | 247.3 | 0 | 0 |
| Hou Zhenzi site | 1001.5 | 14.0 | 120.4 | 0 | 200.0 |
| Hua Erping site | 0 | 0 | 10.3 | 0 | 0 |
| Chang Ping site | 0 | 0 | 202.9 | 0 | 0 |
| HT site | 0 | 0 | 140.0 | 0 | 0 |
| Ban Fangzi site | 0 | 0 | 233.1 | 0 | 0 |
| Dong site | 0 | 0 | 19.0 | 0 | 0 |
| Yu Dongquan site | 0 | 20.0 | 65.9 | 0.5 | 0 |
Table 5.
Environmental factors in the study area.
| Site | T (℃) | pH | A (m) | Dep (m) | H'm | H'o | Tur | DO (mg/L) | F(m/s) |
| WangJia | 18.93 | 8.37 | 978 | 0.05 | 1.966 | 0.5623 | 3.4 | 6.96 | 0.1934 |
| Dong | 19.05 | 8.29 | 1000 | 0.12 | 1.640 | 1.3320 | 3.4 | 6.88 | 0.1593 |
| Ba Muping | 18.22 | 8.63 | 1230 | 1.12 | 0.931 | 1.2150 | 4.0 | 7.22 | 0.1215 |
| Da Manggou | 17.14 | 8.69 | 1251 | 0.46 | 1.661 | 1.4750 | 3.5 | 8.40 | 0.6548 |
| Hu Bao | 21.50 | 8.19 | 888 | 0.07 | 1.869 | 0.1047 | 2.8 | 6.82 | 0.1841 |
| HH | 21.87 | 8.36 | 802 | 0.98 | 1.257 | 1.3800 | 5.8 | 6.71 | 0.4122 |
| Ban Fangzi | 21.00 | 8.53 | 1074 | 0.41 | 2.273 | 0.6365 | 2.8 | 7.86 | 0.3744 |
| Chang Ping | 19.27 | 8.60 | 1141 | 0.05 | 2.312 | 1.1350 | 4.6 | 7.64 | 0.1674 |
| Hou Zhenzi | 16.70 | 8.43 | 1264 | 1.25 | 1.72 | 1.1630 | 3.9 | 7.46 | 0.1433 |
| HT | 20.31 | 8.53 | 1146 | 0.45 | 1.865 | 1.8360 | 5.7 | 7.78 | 0.4923 |
| Chen Jiagou | 17.28 | 8.46 | 645 | 0.23 | 1.382 | 0.6721 | 3.6 | 8.34 | 0.3053 |
| Yu Dongquan | 20.23 | 8.36 | 933 | 1.37 | 1.306 | 1.6420 | 4.7 | 7.22 | 0.1014 |
| Hua Erping | 20.76 | 8.78 | 1321 | 1.32 | 2.019 | 1.3860 | 3.9 | 6.65 | 0.1092 |
| HD | 18.98 | 8.34 | 1032 | 1.45 | 1.427 | 0.1914 | 3.7 | 7.49 | 0.1258 |
Note: T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.
Table 6.
Correlations between biomass of B. lenok tsinlingensis and environmental factors.
| Biomass | T | pH | A | Dep | H'm | H'o | Tur | DO | F | |
| Biomass | 1 | |||||||||
| T | -0.0498 | 1 | ||||||||
| pH | -0.0486 | -0.2220 | 1 | |||||||
| A | -0.0220 | -0.2166 | 0.6626 | 1 | ||||||
| Dep | 0.5700 | -0.0084 | 0.1815 | 0.3115 | 1 | |||||
| H'm | -0.1644 | 0.2324 | 0.1840 | 0.2770 | -0.4935 | 1 | ||||
| H'o | -0.2934 | -0.0304 | 0.4616 | 0.3712 | 0.2307 | -0.1529 | 1 | |||
| Tur | 0.0179 | 0.2417 | 0.1218 | -0.0230 | 0.2963 | -0.2613 | 0.6731 | 1 | ||
| DO | 0.0621 | -0.5661 | 0.3360 | 0.0064 | -0.2143 | 0.0686 | 0.0327 | -0.1016 | 1 | |
| F | -0.3184 | -0.0283 | 0.2593 | -0.0182 | -0.3133 | 0.0910 | 0.2885 | 0.1993 | 0.5753 | 1 |
Note: T, Water temperature; A, Altitude of the sampling site; Dep, Water depth at the sampling point; H'm, Shannon-Weiner index of Benthic Macroinvertebrates; H'o, Shannon-Weiner index of Phytoplankton; Tur, Turbidity; DO, Dissolved Oxygen; F, Flow velocity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



