Submitted:
29 August 2025
Posted:
01 September 2025
You are already at the latest version
Abstract
Canola (Brassica napus L.) is a valuable oilseed plant with high agronomic and industrial value globally, but its genetic improvement is typically constrained by low regeneration efficiency and strong genotype dependence. Advances in tissue culture technology have brought new strategies for overcoming such limitations through realization of developmental plasticity of diverse explants and optimization of hormonal regimes. Current studies show that hypocotyls, cotyledons, petioles, roots, and microspores exhibit extremely variable regeneration responses, the best of which are cotyledons in optimized cytokinin–auxin combinations. In addition, transient culture on hormone-free Murashige and Skoog (MS) medium, or a so-called "shock" treatment, has been shown to be a successful technique of endogenous hormone balance resetting, enhancement of rooting, and mimicking natural withdrawal phases favorable for organogenic reprogramming. Mechanistic insight from Arabidopsis thaliana recognizes the epigenetic hurdles, such as Polycomb Repressive Complex 2 (PRC2), to suppress embryogenic competence, while transient exposure to auxin and after hormone withdrawal facilitates transcriptional activation of crucial regulators such as WUSCHEL (WUS), BABY BOOM (BBM), and LEAFY COTYLEDON (LEC). The synthesis of these findings with Brassica tissue culture offers new opportunities for optimization of the regeneration protocol using a mix of hormonal and epigenetic modulation. New biotechnological applications like hairy root transformation, rol gene use, and somatic embryogenesis-mediated induction of metabolites in similar medicinal plants further extend the application of these technologies. Collectively, these advances provide the foundation for improving more efficient regeneration systems in canola, rationalizing transformation pipelines, and enabling new routes to crop modification and metabolic engineering.
Keywords:
Brassica napus L.
; canola
; tissue culture
; regeneration
; explant response
; hormone-free shock treatment
; somatic embryogenesis
; doubled haploids
; CRISPR/Cas9
; nanotechnology
; bioreactors
; metabolomics
; plant biotechnology
1. Introduction
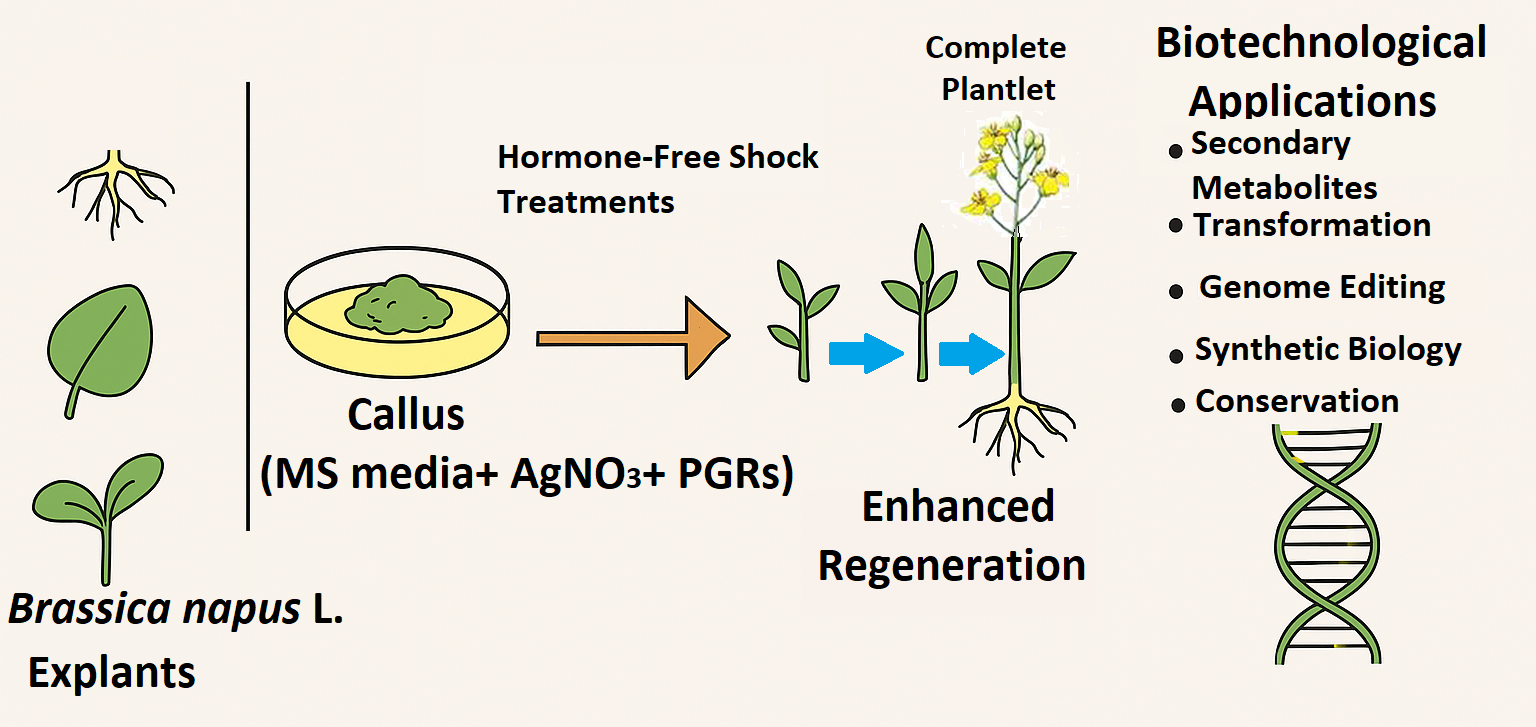
Canola (Brassica napus L.) is a global oilseed crop, and its value encompasses vegetable oil, animal nutrition, therapeutics and renewable energy. Its significance is also growing with rising demand for healthier oils i.e., fats, proteins, biofuels and agricultural climate change resilience (Lin et al., 2013; Goyal et al., 2020; So et al., 2021; Tileuberdi et al., 2022; Yirmibeş & Ülger, 2024; Dastres et al., 2025)). To meet these challenges, plant tissue culture is a foundation for clonal multiplication, gene transformation, and breeding technology. However, B. napus in vitro regeneration is highly genotype-dependent and largely limited by low regeneration efficiency (Naz et al., 2018; Farooq et al., 2019; Dina et al., 2019; Ferrie & Polowick, 2022; Toma and Islam, 2023; Pathirana, 2024).
Explant type (cotyledons, hypocotyls, leaves, embryos) and Plant growth regulators (PGRs)/hormonal regimes (auxins and cytokinins) are the pivotal factors in callogenesis, shoot organogenesis, and root induction (Lone et al., 2016) (Figure 1). Recently, a very promising technique brief culturing on hormone-free MS medium (“shock treatments”) has evidenced enhanced efficiency of rooting and regeneration in some cultivars. These techniques coupled with doubled haploid technology, somatic embryogenesis, and gene-editing systems are revolutionizing tissue culture of B. napus (Khan et al., 2010; Gunjan et al., 2013; Rajesh et al., 2014; Khan et al., 2024). Mozgova et al., (2017) discovered that wild type Zygotic embryos which were cultured on hormone-free medium for an extra 7 days had accelerated the growth of mature primary somatic embryos.
This review critically assesses progress in explant responses, shock treatments independent of Plant growth regulators (PGRs), and new applications, with emphasis on bridging traditional in vitro approaches with biotechnology-induced innovations.
2. Methodology
The present review was conducted by a systematic search of literature to collect, analyze, and synthesize research on tissue culture and regeneration of Brassica napus L. The process underwent a few steps:
- Literature Search Strategy
A comprehensive search was made in premier scientific databases like Scopus, PubMed, and Google Scholar between the years 2010-2025, and occasionally including some earlier landmark papers that established primary tissue culture protocols in B. napus. The keywords used were:
- “Brassica napus tissue culture”
- “canola regeneration”
- “explant responses in Brassica”
- “hormone-free MS medium” OR “shock treatment in plant regeneration”
- “somatic embryogenesis in canola”
- “doubled haploids Brassica napus”
- “genetic transformation canola”
- “CRISPR Brassica napus tissue culture”
- “nanotechnology in plant regeneration”
Boolean operators (AND, OR) were used to limit searches and recover studies that used both traditional tissue culture and new biotechnological uses.
- 2.
- Inclusion and Exclusion Criteria
To maintain quality and relevancy, the following inclusion criteria were used:
-
Inclusion criteria:
- ○
- Peer-reviewed original research papers, reviews, theses, and conference proceedings describing Brassica napus tissue culture.
- ○
- Experiments involving data on explant type response, hormonal treatment, hormone-free “shock” treatments, and biotechnological applications.
- ○
- Regeneration protocols of B. napus experiments.
- ○
- Research on genotype variation and explant-specific response.
- ○
- Hormone-free shock treatment or modified MS protocol reports.
- ○
- Applications of new biotechnology (e.g., doubled haploids, gene editing, metabolomics, and nanotechnology) to canola tissue culture.
- ○
- English-language publications.
-
Exclusion criteria:
- ○
- Literature covering only Brassica species other than B. napus unless directly comparable or as supporting evidence.
- ○
- Papers limited to agronomic field trials with no in vitro regeneration components.
- ○
- Non-peer-reviewed literature before or after year 2012, except where critical historical or methodological significance is involved.
- 3.
- Data Extraction and Organization
Relevant data were systematically scraped and grouped under the following categories:
- Types of explants and regenerability.
- Combinations of hormonal regimes.
- Hormone-free shock treatments and perceived benefits.
- Somatic embryogenesis and doubled haploid methodologies.
- Integration of cutting-edge technologies (CRISPR, nanotechnology, metabolomics, and bioreactors).
All studies were examined for the source of explant, genotype utilized, composition of the culture medium, concentration of Plant growth regulators (PGRs), regeneration efficiency, and results of innovative treatments.
- 4.
- Data Synthesis
Recovered data were cross-matched and tabulated in readable summary tables to facilitate interpretation. Data were combined into comparative tables of hormone-free treatment and explant response to define reproducible patterns and outcomes. Tables highlight similarities and contrasts between protocols, genotype-specific responses, and regeneration effectiveness with varying treatments. New technologies were screened for feasibility, worth, and potential complementarity to standard regeneration protocols.
- 5.
- Quality Assessment
Studies were evaluated on the basis of sample size, replicability of results, description of methods, and statistical support of outcome results.Quantitative regeneration efficiency was preferred more over qualitative descriptive outcomes.
- 6.
- Review Structure
The results were organized into thematic sections, starting with traditional tissue culture and regeneration responses, progressing to PGRs and PGRs/hormone-free shock treatments, and ending with the inclusion of advanced biotechnological instruments. This order was taken to provide both historical evolution and present trends in canola tissue culture studies.
3. Results
3.1. Explant Responses in Brassica napus
Explant type is the most important regeneration determinant. Hypocotyls and cotyledons continue to be the most commonly utilized because of their high responsiveness. Immature embryos and leaf discs have also been used, though success is genotype dependent (Alam et al., 2014).
The findings (Table 1) show that out of the different explants screened for shoot regeneration in Brassica napus, cotyledons proved to be the most uniform and efficient, giving the highest mean shoot number per explant and achieving regeneration efficiencies of up to 100% under optimized conditions (Farooq et al., 2019; Dina et al., 2019; Lone et al., 2016; Biswas et al., 2017). Hypocotyls were fairly effective, producing a mean of 3.3 shoots per explant, with genotype-dependent performance varying from nearly zero to relatively high success in well-suited cultivars (Farooq et al., 2019; Maheshwari et al., 2011; Dina et al., 2019; Ahmad et al., 2016). Petioles regenerated with less capacity compared to cotyledons and hypocotyls, yet with variable success up to 79.3% regeneration in some examples (Farooq et al., 2019). The weakest regeneration reaction was observed in roots, with very poor shoots per explant and low efficiency, and were not suitable for repeated regeneration (Farooq et al., 2019). Anther culture was intermediate in potential between petiole and hypocotyl, with 40–60% efficiency of regeneration, though it was still highly dependent on genotype and culture regimen (Sayem et al., 2010; Kumari et al., 2015). Generally, cotyledons are the most reliable source of explant for high-frequency regeneration, roots being the least ideal, and the rest intermediate based on genotype and technique.
3.2. PGRs Combinations and Response in Brassica napus
The hormonal information of Brassica napus regeneration (Table 2) indicates differential responses in accordance with the kind and concentration of plant growth regulators used. Use of a low concentration of auxin alone, e.g., 2,4-D at 0.1 mg/L, to hypocotyl explants produced mostly callusing, and 1.4 g of callus was produced, and the response was further improved by adding 5 mg/L silver nitrate (Naz et al., 2018). Conversely, cytokinin-enriched treatments with BAP at 2.5–3 mg/L in hypocotyl explants enhanced higher callus proliferation, producing approximately 2.51 g callus and regeneration efficiencies between 40–52.94%, once more enhanced by the inclusion of silver nitrate (Naz et al., 2018; Alam et al., 2014; Roh et al., 2012). In the use of a higher concentration of BAP (5 mg/L) along with the lower concentration of 2,4-D (0.5 mg/L), cotyledon sections proved to respond maximally, exhibiting up to 100% regeneration, though this remained genotype dependent (Lone et al., 2016). 1.0 mg/L BAP + 0.1 mg/L NAA synergistic cytokinin–auxin combination supported shoot growth from cotyledonary leaf with petiole (63.33%) and hypocotyl (43.33%) within 12–16 days, which indicates the importance of synergistic hormonal interaction towards successful regeneration (Toma and Islam, 2023). Thidiazuron (TDZ) 1.6 mg/L (8 µM) was an effective cytokinin equivalent, yielding 47% shoot regeneration from cotyledons, which was further enhanced with the addition of silver nitrate (8.5 mg/L) (Roh et al., 2012). Hypocotyls were very responsive to the synergy of BAP (2 mg/L) and IAA (0.5 mg/L), achieving 84% regeneration efficacy and yielding a mean of 14.5 shoots per explant, establishing the potential of cytokinin–auxin synergy (Hussain et al., 2014). Lastly, the co-culture of low NAA (0.1 mg/L) with kinetin (0–4 mg/L) demonstrated that both hypocotyls and cotyledons were able to regenerate roots at high efficiency (72% in cotyledons and 81% in hypocotyls), while shoot regeneration remained low (20%), indicating auxin-rich or kinetin-biased conditions are conducive to root induction rather than shoot morphogenesis (Anonymous).
3.3. Hormone-Free “Shock” Treatments
Provisional transfer to hormone-free MS medium significantly improves rooting and survival. Shock phase presumably restarts endogenous hormone ratios and reduces stress induced by prolonged exposure to exogenous regulators as evident from model Arabidopsis thaliana (Nic-Can et al., 2016; Mozgová et al., 2017). Shock Duration on Hormone-Free MS removes the barriers caused by continuous auxin exposure (Table 3).
The information (Table 3) illustrates that in immature zygotic embryos of Arabidopsis thaliana, 2,4-D exposure followed by a 7-day culture on hormone-free MS medium caused effective somatic embryogenesis, as 60–70% of the zygotic embryos developed into somatic embryos. Still, extended culture led the response towards callus-like tissue and sporadic shoot formation. This process was mediated by 2,4-D priming for dedifferentiation, followed by removal that allowed natural IAA gradients to be established, whereas relaxation of PRC2 repression lifted the epigenetic block to allow embryogenesis-related transcription factors to be activated (Mozgová et al., 2017).
In Brassica napus cultivars Cyclone, Star, and Westar, a 7-day temporary hormone-free “shock” treatment following combined 2,4-D (0.5 mg/L) and BAP (0.5 mg/L) exposure enhanced rooting and shoot vigor, particularly when explants were then put on 0.1 mg/L NAA and 2 mg/L BAP. This step imitated the Arabidopsis withdrawal system since hormone-free treatment rejuvenated endogenous auxin–cytokinin gradients with auxin accumulation promoting rooting and augmenting organogenic capacity (Khan et al., 2010; Khan et al., 2024).
In Agrobacterium rhizogenes-transformed hairy roots of Solidago nemoralis, spontaneous regeneration of shoots occurred at low frequency (2–3 shoots per root) within 4–6 weeks, and NAA supplementation enhanced shoot formation further. The process behind this was linked with intracellular reprogramming by the rolC gene on the Ri plasmid that is similar to cytokinin activity and synergizing with natural auxin to develop a hormonal condition favorable for regeneration even if exogenous cytokinins are absent (Gunjan et al., 2013).
In Podophyllum hexandrum Milam ecotype seeds, regeneration occurred by a combined multistage treatment: GA₃ (5 mg/L) stimulated 99.3% germination, 2,4-D (1.5 mg/L) in darkness caused embryogenic callus, and culture transfer to hormone-free MS medium allowed embryo maturation and polarity. This led to extremely high somatic embryogenesis frequencies (89.6% direct SE; 47.7 embryos per 50 mg callus), and 79% of embryos formed normal plants. Additionally, somatic embryos regenerated had much larger amounts of podophyllotoxin (1.8 mg/g DW) than field-grown plants, illustrating how dedifferentiation, stress signaling, and hormone withdrawal promoted embryogenic competence and secondary metabolite production (Rajesh et al., 2014).
3.4. Emerging Biotechnological Applications
- Somatic embryogenesis & synthetic seeds with specific coating material: Permits mass propagation and storage of germplasm (Nongdam, 2016; Iqbal & Möllers, 2019).
- Doubled haploids: Accelerate breeding through homozygous line production (Shmykova et al., 2016; Rahman & de Jiménez, 2016; Starosta et al., 2023).
- CRISPR/Cas9 genome editing: Used for yield improvement (Yang et al., 2018; Zheng et al., 2020), nutritional improvement (Okuzaki et al., 2018; Huang et al., 2020), seed protein improvement (Zhai et al., 2020), oil quality improvement (Xie et al., 2020), stress resistance, and disease resistance (Sun et al., 2018; Li et al., 2022).
- Omics-based optimization: Transcriptomics and metabolomics describe hormone control and stress-response pathways (Zhu et al., 2016; Knoch et al., 2021; Sarkar et al., 2023; Dhiman et al., 2024).
- Biogenic nanoparticles: e.g., Metal or Silver nanoparticle and nanofertilizer cause deposition of metabolites and callogenesis (Ahmed et al., 2023; Assad et al., 2025; Farid et al., 2025).
4. Discussion
The review of literature emphasizes that while traditional regeneration protocols in B. napus are exceedingly genotype-specific, significant progress has been made by optimizing explant selection, hormonal regimes, and the addition of hormone-free shock treatments. Hypocotyl and cotyledon remain popular explants, although doubled haploid approaches via microspores are of crucial importance for breeding (Dina et al., 2019; Farooq et al., 2019).
The physiological rationale for hormone-free shocks is related to re-establishing endogenous auxins and cytokinins, which minimizes callus vitrification and enhances rooting. Such treatments work well for cultivars that were once described as recalcitrant thaliana (Nic-Can et al., 2016; Mozgová et al., 2017; Khan et al., 2010; Khan et al., 2024).
Biotechnology integration has increased applications of tissue culture beyond propagation. CRISPR-Cas gene editing, for instance, demands effective regeneration platforms, and metabolomic profiling informs media optimization. Biogenic nanoparticles and TIBs hold promising frontiers for scalable, sustainable canola improvement (Yang et al., 2018; Zheng et al., 2020; Okuzaki et al., 2018; Huang et al., 2020; Zhai et al., 2020; Xie et al., 2020; Sun et al., 2018; Li et al., 2022).
4.1. Explant-Specific Variation in Regeneration Competence of Brassica napus
The comparative regenerations of Brassica napus explants (Table 1) clearly show that regeneration efficiency is highly explant-dependent and highly genotype and culture protocol-dependent (Farooq et al., 2019). The best explant found was cotyledons, with a mean value of 9 shoots/explant, a figure not far from three times that of hypocotyls (3.3 shoots/explant) and more than three times that of petioles (2.8 shoots/explant). Reported percentages of regeneration for cotyledons vary from 6% to 100% and confirm that even though they can regenerate completely under optimal protocols, their regeneration can be heterogeneous between cultivars, perhaps due to the fact that they possess actively dividing, juvenile cells with high meristematic potential. Being storage organs in the seed, they are metabolically loaded with nutrients and readily respond to exogenous growth regulators such as BAP and NAA. They also contain more flexible and less developmentally restricted cells than mature tissue, which contributes to their competence for shoot organogenesis (Farooq et al., 2019; Dina et al., 2019; Lone et al., 2016; Biswas et al., 2017). Hypocotyls, although less effective than cotyledons, were also of moderate competence, with efficiencies varying from 0% to 50.6% in some and 82% in others, indicating that specific genotypes or specially optimized protocols can liberate much greater regeneration frequencies than general values predict, are very efficient since they consist of long parenchymatous cells that can dedifferentiate upon auxin–cytokinin treatment. Yet, their regenerative response is highly genotypic dependent, perhaps due to variations in endogenous hormone concentration and gene expression patterns governing shoot meristem identity. (Farooq et al., 2019; Maheshwari et al., 2011; Dina et al., 2019; Ahmad et al., 2016). Petioles were less reliable, producing on average only 2.8 shoots per explant with efficiencies ranging from 4% to 79.3%, indicating that they are responsive but more unpredictable in their performance (than either hypocotyls or cotyledons), are less successfully regenerated since their cells are specialized and structurally differentiated with little meristematic potential. (Farooq et al., 2019). Roots were the least efficient explant, averaging below 2.35 shoots per explant and rarely higher than 42.6% efficiency, and thus would not be good choices for efficient regeneration investigations, are highly specialized during the process of absorption and possess stronger differentiation patterns. Endogenous auxin at higher concentrations is also present in roots, which may inhibit shoot organogenesis unless contained by cytokinins (Farooq et al., 2019). Anther culture provided another perception, with approximately 2.44 shoots per callus with efficiencies of 40–60%, thus outmatching roots but being excessively dependent on the genotype (Sayem et al., 2010; Kumari et al., 2015). Overall, these figures highlight that cotyledons are the qualitative superior explant for regeneration of canola with the yield reaching up to 100% efficiency while roots provide the poorest response, highlighting the importance of explant selection in tissue culture improvement.
4.2. Hormonal Regulation and Molecular Determinants of Explant Regeneration in Brassica napus
The auxin- and cytokinin-regulated reactions of Brassica napus regeneration (Table 2) are understood in relation to the control by auxin and cytokinin of cellular reprogramming and organogenic balance. Low concentrations of auxin alone, such as 2,4-D at 0.1 mg/L, largely induced dedifferentiation, leading to callus formation at the expense of organized shoots as auxin promotes cell proliferation but suppresses shoot organogenesis when used alone (Naz et al., 2018). When cytokinin in the form of BAP was applied at intermediate levels (2.5–3 mg/L), hypocotyl explants formed higher callus biomass with incomplete regeneration efficiency since cytokinins induced some shoot primordia but required either auxin synergy or further signals like silver nitrate to activate shoot formation pathways to completion (Naz et al., 2018; Alam et al., 2014; Roh et al., 2012). The extremely potent 5 mg/L BAP + 0.5 mg/L 2,4-D combination in cotyledons implies that an incidentally delivered pulse of auxin in combination with intense cytokinin stimulation sends the appropriate hormonal signals to initiate meristematic regulators, and this is why virtually 100% regeneration was achieved here, albeit genetic background still remained the determining factor (Lone et al., 2016). Also, a balanced mixture of 1 mg/L BAP with 0.1 mg/L NAA induced cotyledonary and hypocotyl explants to produce shoots in 12–16 days, proving that low auxin levels cooperate with moderate cytokinin to induce organized shoot meristems optimally (Toma and Islam, 2023). Thidiazuron, being a potent cytokinin analog, mimicked the effects and caused almost 50% regeneration of shoots from cotyledons, reflecting its major contribution to shoot morphogenesis (Roh et al., 2012). The IAA plus BAP combination was particularly superior, with 84% regeneration in hypocotyls and giving a mean of 14.5 shoots per explant, indicating that IAA, in contrast to synthetic auxins 2,4-D or NAA, offers a more natural gradient to promote meristem organization (Hussain et al.,2014). In contrast, NAA application in combination with kinetin biased regeneration toward root rather than shoot formation, with hypocotyls attaining 81% rooting compared to a mere 20% shoot induction, indicative of the auxin-prepondowering condition predisposing toward root over shoot organogenesis. These observations collectively establish that shoot regeneration in B. napus is subject to strict cytokinin–auxin balance, with cytokinins being the most important factor but auxins playing a critical modulating role on cellular competence and organ identity (Anonymous).
4.3. Molecular Determinants of Explant Regeneration in Brassica napus
The hormone-specific responses in Brassica napus regeneration can be traced back to the activation or repression of central molecular regulators that determine embryogenic competence and meristem identity (Figure 2). At low levels of 2,4-D treatment, dedifferentiation is favored but without strong cytokinin signaling, transcription factors like WUSCHEL (WUS) that ensures shoot apical meristem (SAM) identity and SHOOT MERISTEMLESS (STM) that ensures meristematic cell proliferation are not adequately induced and thus result in callus growth instead of organized shoot development. On the other hand, treatments with predominant BAP or TDZ give intense cytokinin signals triggering WUS and CUP-SHAPED COTYLEDON (CUC) genes that are important in the shoot meristem initiation and thus the reason why intermediate to high levels of BAP and auxin pulses lead to high-frequency shoot regeneration (Elhiti & Stasolla, 2012).
In Arabidopsis LATERAL ORGAN BOUNDARIES DOMAIN (LBD)/ASYMMETRIC LEAVES2-LIKE (ASL) transcription factors play a role in regulating callus formation program. The four LBD ARF-downstream genes, LBD16, LBD17, LBD18 and LBD29, are dramatically and quickly induced by callus-inducing medium (CIM) in various organs (Fan et al., 2012; Ikeuchi et al., 2013).
The very effective co-action of 5 mg/L BAP and 0.5 mg/L 2,4-D in cotyledons most probably acts by conditionally activating the BABY BOOM (BBM) and LEAFY COTYLEDON (LEC1/LEC2) genes that reprogram somatic cells for embryogenic identity and cytokinin dominance for WUS-mediated formation of shoot meristems. Likewise, BAP + NAA treatment reveals that reduced levels of auxin not only counterbalance cytokinin action but also aid in the expression of AUXIN RESPONSE FACTORS (ARFs), which contribute to the formation of local auxin maxima required for organ primordia initiation. The BAP + IAA treatment accentuates this even more, as IAA, the natural auxin, complements cytokinin signaling to activate WUS and STM more effectively and also facilitates the co-activation of BBM and LEC genes that account for the resultant high shoot numbers per explant (Ikeuchi et al.,2016).
In contrast, if NAA is co-cultivated with kinetin, the endogenous hormonal status is skewed toward auxin predominance, and this encourages root identity by activating ROOT APICAL MERISTEM (RAM) regulators such as PLETHORA (PLT) genes and inhibiting shoot meristem regulators like WUS. This can account for rooting frequencies exceeding 80% and shoot regeneration frequencies below 20% (Iwase et al., 2018). Therefore, the data show that effective regeneration is not a question of hormonal balance, but how these hormone signals are processed at the molecular level by major transcriptional networks. In canola, as in Arabidopsis, whether a callus develops into a root, a shoot, or an embryo is determined by the capacity to modulate expression of WUS, BBM, LEC, and PLT genes under given auxin–cytokinin conditions (Ikeuchi et al., 2019; Shen et al., 2020).
4.4. Hormone Withdrawal, Epigenetic Reprogramming, and Biotechnological Applications of Somatic Embryogenesis Across Plant Systems
The comparative data (Table 3, Figure 3) indicate that regeneration in Arabidopsis thaliana, Brassica napus, Solidago nemoralis, and Podophyllum hexandrum all point to a unifying principle: temporary hormone-free periods (or internal hormonal reprogramming) provide a permissive context for dedifferentiated cells to recover totipotency and reactivate developmental programs (Garcia, 2019). Yet molecular regulators, hormone sensitivities, and biotechnological rationales vary among species. Arabidopsis has the mechanistic model (epigenetic barrier release through PRC2 relaxation), Brassica shows agricultural significance (enhanced cultivar regeneration for transformation), Solidago exhibits endogenous hormone-mimetic engineering (rolC), and Podophyllum reveals the connection between somatic embryogenesis and secondary metabolism (podophyllotoxin production) (Khan et al., 2010; Rajesh et al., 2014; Gunjan et al., 2013; Mozgová et al., 2017; Khan et al., 2024).
In Arabidopsis thaliana, 2,4-D priming with subsequent hormone withdrawal triggered somatic embryogenesis by permitting the development of indole-3-acetic acid (IAA) gradients and relieving PRC2-mediated chromatin repression. PRC2 typically lays down H3K27me3 marks on important embryogenic regulators like LEAFY COTYLEDON1/2 (LEC1/2), BABY BOOM (BBM), and WUSCHEL (WUS). When PRC2 activity is diminished, either genetically or through culture-induced stress, such transcription factors are derepressed, facilitating reprogramming of somatic cells to embryogenic states. This points to a fundamental molecular hurdle in plant totipotency: epigenetic silencing of embryo regulators. The biotechnology significance is that alteration of PRC2 activity, by epigenetic editing or reversible chemical inhibition, would greatly enhance somatic embryogenesis protocols in recalcitrant crops (Mozgová et al., 2017).
In Brassica napus, a close relative crop, comparable hormone-free shocks following exposure to auxin–cytokinins enhanced shoot vigor and rooting. This replicates the Arabidopsis model in which transient auxin treatment primes dedifferentiation and withdrawal permits endogenous hormone rebalancing and competence for organogenesis. Molecularly, the cytokinins from BAP would have activated WUS and CUP-SHAPED COTYLEDON (CUC) to specify shoot meristem identity, and transient auxin exposure assisted ARF-mediated local auxin maxima for primordium initiation. Shock phase is found to reestablish endogenous hormonal crosstalk, promoting rooting via PLETHORA (PLT) induction while ensuring shoot regeneration. From a biotechnological perspective, such a strategy illustrates how the control of temporal exposure to Plant growth regulators (PGRs) enhances regeneration efficacy in elite cultivars and speeds up transformation pipelines for canola improvement (Khan et al., 2010; Khan et al., 2024).
In Solidago hemoralis, regeneration from hairy roots depended less on withdrawal of exogenous hormone and more on reprogramming endogenously by rolC gene brought in by Agrobacterium rhizogenes. The rolC gene produces a protein that simulates endogenous cytokinin signaling, modifying the ratio of auxin to cytokinin within cells. Combined with endogenous auxin, this favored low-frequency shoot regeneration even without cytokinins. This example demonstrates how genetic engineering using Ri plasmid genes can overcome exogenous hormone needs, albeit at reduced efficiencies. Biotechnologically, rol genes offer a means for the induction of morphogenic competence, which may be of use in species where tissue culture is strongly recalcitrant (Gunjan et al., 2013).
In Podophyllum hexandrum, an endangered medicinal plant subjected to overharvesting, the regeneration system comprised a multistep protocol: GA₃ induced seed dormancy break and enhanced germination, 2,4-D caused embryogenic callus, and hormone-free culture facilitated somatic embryo maturation and polarity. Significantly, this system not only yielded high-frequency somatic embryos (89.6%) but also increased secondary metabolite production, somatic embryos contained 1.8 mg/g DW podophyllotoxin, which was much higher than field-grown plants. This observation discloses somatic embryogenesis not only as a tool of propagation but as well as a biotechnological approach to metabolite improvement, since dedifferentiated and reprogrammed cells tend to induce stress-related and secondary metabolism pathways. At the molecular level, this could be through crosstalk between embryogenesis regulators (LEC, BBM) and metabolic pathway genes by virtue of common hormone signaling networks (e.g., auxin-responsive transcription factors controlling phenylpropanoid pathways) (Rajesh et al., 2014).
Cumulatively, these findings show that although the hormone withdrawal or shock principle is generally applicable to plant regeneration, the responses are species-specific and dependent on genetic/epigenetic context, endogenous hormone dynamics, and even transgene influence. Contemporary biotechnological strategies like CRISPR-based epigenome editing of PRC2 targets, synthetic auxin/cytokinin delivery systems, or rol gene engineering may render such regeneration methods more predictable and efficient. In addition, integration of somatic embryogenesis with metabolic engineering confers a double benefit: propagation of unusual plants at rapid rates and improved yields of useful metabolites such as podophyllotoxin (Khan et al., 2010; Rajesh et al., 2014; Gunjan et al., 2013; Mozgová et al., 2017; Khan et al., 2024).
5. Future Prospects
Tissue culture and Brassica napus regeneration are founded on the development of genotype-independent, universal protocols with effective applicability on a series of cultivars. The integrated use of omics-enabled fields such as transcriptomics, proteomics, and metabolomics will reveal the molecular processes of regeneration, enabling the development of ideal media and culture conditions.Genome editing technologies such as CRISPR/Cas9 will increasingly rely on efficient regeneration systems, and refinement of explant-specific responses becomes even more important. New technologies such as nanomaterials, temporary immersion bioreactors, and synthetic seed production are likely to boost callus induction, bulk propagation, and germplasm conservation. Artificial intelligence and machine learning models could soon be used to forecast regeneration success based on genotype and culture parameters, minimizing trial-and-error experimentation. All these developments combined will make canola tissue culture a highly productive platform for crop enhancement, bioengineering, and green agriculture.
6. Conclusions
The Brassica napus regeneration has been highly developed with optimization of explant selection, hormone-free shock treatment, and implementation of novel biotechnological equipment, but genotype specificity and heterogeneity of protocols are persistent problems. Future advances will depend on the integration of conventional tissue culture with systems biology expertise, genome editing, nanotechnology, and large-scale propagation systems for creating more universal, efficient, and reproducible regeneration platforms. Through the combination of metabolomics-assisted optimization, machine learning-driven prediction, and synthetic seed technology, canola tissue culture can become a state-of-the-art system that not only facilitates breeding and genetic transformation but also expedites the production of stress-tolerant, nutritionally enriched, and industrially improved cultivars for the sustenance of the crop’s place in global food security and sustainable agriculture. In spite of progress, challenges are still there: genotype-dependency, somaclonal variation, and labor-intensive protocols. Overcoming these challenges necessitates integration of traditional tissue culture with systems biology, nanotechnology, and machine learning-based predictive models for regeneration success.
Acknowledgment
We are very much thankful to Dr. Farooq, Agriculture Research Institute, University of Agriculture, North, Mingora, Khyber Pukhtunkhwa, Pakistan. Special thanks to Shah Meer Ali Khan (BS Computer Science, National College of Business Administration & Economics, Lahore, Punjab, Pakistan)
Conflict of Interest
The authors declare no conflict of interest.
References
- Ahmad, K., Jalal, A., Rajab, H., Ullah, M., & Khan, M. S. (2016). Screening of promising Brassica napus L. genotypes for callus induction and regeneration. International Journal of Biology and Biotechnology, 13, 203-215. https://ijbbku.com/assets/custom/journals/2016/2/Screening%20of%20promising%20Brassica%20napus%20L.%20Genotypes%20for%20callus%20induction%20and%20regeneration.pdf.
- Ahmed, T., Masood, H. A., Noman, M., Al-Huqail, A. A., Alghanem, S. M., Khan, M. M., Muhammad, S., Manzoor, N., Rizwan, M., Qi, X. and Abeed, A.H., & Li, B. (2023). Biogenic silicon nanoparticles mitigate cadmium (Cd) toxicity in rapeseed (Brassica napus L.) by modulating the cellular oxidative stress metabolism and reducing Cd translocation. Journal of Hazardous Materials, 459, 132070. https://www.sciencedirect.com/science/article/abs/pii/S0304389423013535.
- Alam, S. S., Khaleda, L., & Al-Forkan, M. (2014). In vitro plant regeneration from hypocotyl and cotyledon explants of bari sarisha-13 (Brassica napus L.). Asian Journal of Agriculture and Food Sciences, 2(1). https://www.researchgate.net/profile/Laila-Khaleda/publication/347973680_In_vitro_Plant_Regeneration_from_Hypocotyl_and_Cotyledon_Explants_of_BARI_SARISHA-13_Brassica_napus_L/links/5feaddfe92851c13fed0030d/In-vitro-Plant-Regeneration-from-Hypocotyl-and-Cotyledon-Explants-of-BARI-SARISHA-13-Brassica-napus-L.pdf.
- Assad, N., Ansa, Laila, M. B., Ullah, S., Adnan, M., Wahab, S., Kaplan, A., Khan, M.N., & Ullah, B. (2025). Nanotechnology in Oilseed Crops: A New Frontier for Abiotic Stress Adaptation. In Oilseed Crops Under Abiotic Stress: Mitigation Strategies and Future Perspectives (pp. 507-536). Singapore: Springer Nature Singapore. https://link.springer.com/chapter/10.1007/978-981-96-8346-8_16.
- Biswas, S. K., Tareq, M. Z., Ahmmed, S., Hoque, A. B. M. Z., & Rahman, M. T. (2017). Callus derived regeneration of some selected Brassica genotypes. The Agriculturists, 15(2), 1-10. https://banglajol.info/index.php/AGRIC/article/view/35458.
- Dastres, E., Sarvestani, G. S., Edalat, M., & Pourghasemi, H. R. (2025). Habitat suitability modeling of dominant weed in canola (Brassica napus) fields using machine learning techniques. Weed Science, 73, e35. https://www.cambridge.org/core/journals/weed-science/article/habitat-suitability-modeling-of-dominant-weed-in-canola-brassica-napus-fields-using-machine-learning-techniques/CD0C79F13552E16798A3853B1DF297E1.
- Dhiman, V. K., Singh, D., Dhiman, V. K., & Pandey, H. (2024). Multi-omics: An Advanced Bioinformatics Approach for Crop Improvement in Agriculture. Bioinformatics for Plant Research and Crop Breeding, 75-98. https://onlinelibrary.wiley.com/doi/abs/10.1002/9781394209965.ch3.
- Dina, M. M. A., Sultana, S., & Bhuiyan, M. S. U. (2019). Development of high frequency in vitro plant regeneration protocol of Brassica napus. Journal of Advanced Biotechnology and Experimental Therapeutics, 2(3), 114-119. https://www.researchgate.net/profile/Sayeda-Sultana/publication/335974799_Developing_high_frequency_in_vitro_plant_regeneration_protocol_of_Brassica_napus/links/6375c8d654eb5f547cdc132a/Developing-high-frequency-in-vitro-plant-regeneration-protocol-of-Brassica-napus.pdf?origin=journalDetail&_tp=eyJwYWdlIjoiam91cm5hbERldGFpbCJ9.
- Elhiti, M., & Stasolla, C. (2012). In vitro shoot organogenesis and hormone response are affected by the altered levels of Brassica napus meristem genes. Plant Science, 190, 40-51. https://www.sciencedirect.com/science/article/abs/pii/S0168945212000660.
- Fan, M., Xu, C., Xu, K., & Hu, Y. (2012). LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation in Arabidopsis regeneration. Cell research, 22(7), 1169-1180. https://www.nature.com/articles/cr201263.
- Farid, M., Zulfiqar, A., Farid, S., Alshehri, M. A., Alomrani, S. O., Asam, Z. U. Z., Sarfraz, W., Ghafoor, A., & Ali, S. (2025). Combined application of Azadirachta indica leaf extract and zerovalent iron nanoparticles (nZVI) enhances phytoremediation potential of Brassica napus L. in surgical industry wastewater. International Journal of Phytoremediation, 1-13. https://www.tandfonline.com/doi/abs/10.1080/15226514.2025.2537201.
- Farooq, N., Nawaz, M. A., Mukhtar, Z., Ali, I., Hundleby, P., & Ahmad, N. (2019). Investigating the in vitro regeneration potential of commercial cultivars of Brassica. Plants, 8(12), 558. https://www.mdpi.com/2223-7747/8/12/558.
- Farooq, N., Nawaz, M. A., Mukhtar, Z., Ali, I., Hundleby, P., & Ahmad, N. (2019). Investigating the in vitro regeneration potential of commercial cultivars of Brassica. Plants, 8(12), 558. https://www.mdpi.com/2223-7747/8/12/558.
- Garcia, C., Furtado de Almeida, A. A., Costa, M., Britto, D., Valle, R., Royaert, S., & Marelli, J. P. (2019). Abnormalities in somatic embryogenesis caused by 2, 4-D: an overview. Plant Cell, Tissue and Organ Culture (PCTOC), 137(2), 193-212. https://link.springer.com/article/10.1007/s11240-019-01569-8.
- Goyal, A., Tanwar, B., Sihag, M. K., Kumar, V., Sharma, V., & Soni, S. (2020). Rapeseed/canola (Brassica napus) seed. In Oilseeds: Health Attributes and Food Applications (pp. 47-71). Singapore: Springer Singapore. https://link.springer.com/chapter/10.1007/978-981-15-4194-0_2.
- Gunjan, S. K., Lutz, J., Bushong, A., Rogers, D. T., & Littleton, J. (2013). Hairy root cultures and plant regeneration in Solidago nemoralis transformed with Agrobacterium rhizogenes. American Journal of Plant Sciences, 2013. https://www.scirp.org/html/18-2600917_35477.htm.
- Huang, H., Cui, T., Zhang, L., Yang, Q., Yang, Y., Xie, K., Fan, C., & Zhou, Y. (2020). Modifications of fatty acid profile through targeted mutation at BnaFAD2 gene with CRISPR/Cas9-mediated gene editing in Brassica napus. Theoretical and Applied Genetics, 133(8), 2401-2411. https://link.springer.com/article/10.1007/s00122-020-03607-y.
- Hussain, S., Rasheed, A., Latif, M., Mahmood, T. A. R. I. Q., & Naqvi, S. S. (2014). Canola (Brassica napus L.) regeneration and transformation via hypocotyl and hypocotyl derived calli. Sarhad J. Agric, 30(2), 165-172. https://www.researchgate.net/profile/Saira-Hussain-5/publication/282664483_CANOLA_BRASSICA_NAPUS_L_REGENERATION_AND_TRANSFORMATION_VIA_HYPOCOTYL_AND_HYPOCOTYL_DERIVED_CALLI/links/5617605108ae839f3c7d8956/CANOLA-BRASSICA-NAPUS-L-REGENERATION-AND-TRANSFORMATION-VIA-HYPOCOTYL-AND-HYPOCOTYL-DERIVED-CALLI.pdf.
- Ikeuchi, M., Favero, D. S., Sakamoto, Y., Iwase, A., Coleman, D., Rymen, B., & Sugimoto, K. (2019). Molecular mechanisms of plant regeneration. Annual review of plant biology, 70(1), 377-406. https://www.annualreviews.org/content/journals/10.1146/annurev-arplant-050718-100434.
- Ikeuchi, M., Ogawa, Y., Iwase, A., & Sugimoto, K. (2016). Plant regeneration: cellular origins and molecular mechanisms. Development, 143(9), 1442-1451. https://journals.biologists.com/dev/article/143/9/1442/47902/Plant-regeneration-cellular-origins-and-molecular.
- Ikeuchi, M., Sugimoto, K., & Iwase, A. (2013). Plant callus: mechanisms of induction and repression. The plant cell, 25(9), 3159-3173. https://academic.oup.com/plcell/article/25/9/3159/6097899.
- Iqbal, M. C. M., & Möllers, C. (2019). Towards artificial seeds from microspore derived embryos of Brassica napus. Plant Cell, Tissue and Organ Culture (PCTOC), 139(2), 207-225. https://link.springer.com/article/10.1007/s11240-019-01692-6.
- Iwase, A., Mita, K., Favero, D. S., Mitsuda, N., Sasaki, R., Kobayshi, M., Takebayashi, Y., Kojima, M., Kusano, M., Oikawa, A. and Sakakibara, H., & Sugimoto, K. (2018). WIND1 induces dynamic metabolomic reprogramming during regeneration in Brassica napus. Developmental biology, 442(1), 40-52. https://www.sciencedirect.com/science/article/pii/S0012160618300873.
- Khan, I., Shuaib, M., Khan, S., Zia, M. A., & Akbar, H. (2010). A Micropropagation, Molecular and Physiochemical Analysis of Canola Brassica napus L. From Seeds. Nature Precedings, 1-1. https://www.nature.com/articles/npre.2010.4781.2.
- Khan, I., Ali, W., Akhtar, M. W., & Ahmad, N. (2024). Enhanced rooting and regeneration of canola (Brassica napus L.) cultivars after a brief period (shock) on growth regulator-free Murashige and Skoog (MS) medium. https://academicjournals.org/journal/AJB/article-full-text-pdf/E8AB94072537.pdf.
- Knoch, D., Werner, C. R., Meyer, R. C., Riewe, D., Abbadi, A., Lücke, S., Snowdon, R.J., & Altmann, T. (2021). Multi-omics-based prediction of hybrid performance in canola. Theoretical and Applied Genetics, 134(4), 1147-1165. https://link.springer.com/article/10.1007/s00122-020-03759-x.
- Kumari, P., Singh, A. K., Sharma, M., & Ahmed, S. (2015). Comparative response of different genotypes of brassica to anther culture. International Journal of Research, 3(4), 33-44. https://www.researchgate.net/profile/Preeti-Kumari-13/publication/285734567_Comparative_Response_of_Different_Genotypes_of_Brassica_to_Anther_Culture/links/5662e5eb08ae418a786b8ed9/Comparative-Response-of-Different-Genotypes-of-Brassica-to-Anther-Culture.pdf.
- Li, J., Yu, X., Zhang, C., Li, N., & Zhao, J. (2022). The application of CRISPR/Cas technologies to Brassica crops: current progress and future perspectives. Abiotech, 3(2), 146-161. https://link.springer.com/article/10.1007/s42994-022-00076-3.
- Lin, L., Allemekinders, H., Dansby, A., Campbell, L., Durance-Tod, S., Berger, A., & Jones, P. J. (2013). Evidence of health benefits of canola oil. Nutrition reviews, 71(6), 370-385. https://academic.oup.com/nutritionreviews/article-abstract/71/6/370/1882361.
- Lone, J. A., Gupta, S. K., Wani, S. H., Bhat, M. A., & Lone, R. A. (2016). In vitro regeneration studies in Brassica napus with response to callus induction frequency and regeneration frequency. International Journal of Agriculture, Environment and Biotechnology, 9(5), 755. https://ndpublisher.in/admin/issues/IJAEBV9I5g.pdf.
- Maheshwari, P., Selvaraj, G., & Kovalchuk, I. (2011). Optimization of Brassica napus (canola) explant regeneration for genetic transformation. New biotechnology, 29(1), 144-155. https://www.sciencedirect.com/science/article/abs/pii/S1871678411001567.
- Mozgova, I., Munoz-Viana, R., & Hennig, L. (2017). PRC2 represses hormone-induced somatic embryogenesis in vegetative tissue of Arabidopsis thaliana. PLoS Genetics, 13(1), e1006562. https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1006562.
- Naz, S., Siddiquiland, M. F., & Raza, S. (2018). Effect of different growth regulators on in vitro propagation of Brassica napus L. Pak J Bot, 50(5), 1871-1876. https://pakbs.org/pjbot/papers/1527617060.pdf.
- Nic-Can, G. I., Avilez-Montalvo, J. R., Aviles-Montalvo, R. N., Márquez-López, R. E., Mellado-Mojica, E., Galaz-Ávalos, R. M., & Loyola-Vargas, V. M. (2016). The relationship between stress and somatic embryogenesis. In Somatic embryogenesis: fundamental aspects and applications (pp. 151-170). Cham: Springer International Publishing. https://link.springer.com/chapter/10.1007/978-3-319-33705-0_9.
- Nongdam, P. (2016). Development of synthetic seed technology in plants and its applications: a review. Int J Curr Sci, 19(4), 86-101. https://www.researchgate.net/profile/Potshangbam-Nongdam/publication/343179931_Development_of_Synthetic_Seed_Technology_in_Plants_and_its_Applications_A_Review/links/5f1a7313a6fdcc9626ad33e3/Development-of-Synthetic-Seed-Technology-in-Plants-and-its-Applications-A-Review.pdf.
- Okuzaki, A., Ogawa, T., Koizuka, C., Kaneko, K., Inaba, M., Imamura, J., & Koizuka, N. (2018). CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiology and Biochemistry, 131, 63-69. https://www.sciencedirect.com/science/article/abs/pii/S0981942818301839.
- Pathirana, R. (2024). Tissue Culture Technologies and Their Applications. Combined Proceedings IPPS 74:187-219. https://www.researchgate.net/profile/Ranjith-Pathirana/publication/391089211_Tissue_Culture_Technologies_and_Their_Applications/links/68217980ded4331557468a55/Tissue-Culture-Technologies-and-Their-Applications.pdf.
- Rahman, M., & de Jiménez, M. M. (2016). Behind the scenes of microspore-based double haploid development in Brassica napus: A review. J. Plant Sci. Mol. Breed, 5(1), 1. https://www.researchgate.net/profile/Mukhlesur-Rahman-2/publication/305272664_Behind_the_scenes_of_microspore-based_double_haploid_development_in_Brassica_napus_A_review/links/5a874083458515b8af8d5e0b/Behind-the-scenes-of-microspore-based-double-haploid-development-in-Brassica-napus-A-review.pdf?_sg%5B0%5D=started_experiment_milestone&origin=journalDetail&_rtd=e30%3D.
- Rajesh, M., Sivanandhan, G., Jeyaraj, M., Chackravarthy, R., Manickavasagam, M., Selvaraj, N., & Ganapathi, A. (2014). An efficient in vitro system for somatic embryogenesis and podophyllotoxin production in Podophyllum hexandrum Royle. Protoplasma, 251(5), 1231-1243. https://pubmed.ncbi.nlm.nih.gov/24633328.
- Roh, K. H., Kwak, B. K., Kim, J. B., Lee, K. R., Kim, H. U., & Kim, S. H. (2012). The influence of silver thiosulfate and thidiazuron on shoot regeneration from cotyledon explants of Brassica napus. Journal of Plant Biotechnology, 39(3), 133-139. https://www.researchgate.net/profile/Kyung-Hee-Roh/publication/264058246_The_influence_of_silver_thiosulfate_and_thidiazuron_on_shoot_regeneration_from_cotyledon_explants_of_Brassica_napus/links/53d590f00cf220632f3d5948/The-influence-of-silver-thiosulfate-and-thidiazuron-on-shoot-regeneration-from-cotyledon-explants-of-Brassica-napus.pdf.
- Sarkar, A., Kisiala, A., Adhikary, D., Basu, U., Emery, R. N., Rahman, H., & Kav, N. N. (2023). Silicon ameliorates clubroot responses in canola (Brassica napus): A “multi-omics”-based investigation into possible mechanisms. Physiologia Plantarum, 175(2), e13900. https://onlinelibrary.wiley.com/doi/full/10.1111/ppl.13900.
- Sayem, M. A., Maniruzzaman, M., Siddique, S. S., & Al-Amin, M. (2010). In vitro shoot regeneration through anther culture of Brassica spp. Bangladesh Journal of Agricultural Research, 35(2), 331-341. https://banglajol.info/index.php/BJAR/article/view/5896.
- So, K. K., & Duncan, R. W. (2021). Breeding canola (Brassica napus L.) for protein in feed and food. Plants, 10(10), 2220. https://www.mdpi.com/2223-7747/10/10/2220.
- Shen, S., Sun, F., Zhu, M., Chen, S., Guan, M., Chen, R., Tang, F., Yin, N., Xu, X., Tang, Z. and Li, J., & Qu, C. (2020). Genome-wide identification AINTEGUMENTA-like (AIL) genes in Brassica species and expression patterns during reproductive development in Brassica napus L. Plos one, 15(6), e0234411. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0234411.
- Shmykova, N. A., Shumilina, D. V., & Suprunova, T. P. (2016). Doubled haploid production in Brassica L. species. Russian Journal of Genetics: Applied Research, 6(1), 68-77. https://link.springer.com/article/10.1134/S2079059716010123.
- Starosta, E., Szwarc, J., Niemann, J., Szewczyk, K., & Weigt, D. (2023). Brassica napus haploid and double haploid production and its latest applications. Current Issues in Molecular Biology, 45(5), 4431-4450. https://www.mdpi.com/1467-3045/45/5/282.
- Sun, Q., Lin, L., Liu, D., Wu, D., Fang, Y., Wu, J., & Wang, Y. (2018). CRISPR/Cas9-mediated multiplex genome editing of the BnWRKY11 and BnWRKY70 genes in Brassica napus L. International journal of molecular sciences, 19(9), 2716. https://www.mdpi.com/1422-0067/19/9/2716.
- Tileuberdi, N., Turgumbayeva, A., Yeskaliyeva, B., Sarsenova, L., & Issayeva, R. (2022). Extraction, isolation of bioactive compounds and therapeutic potential of rapeseed (Brassica napus L.). Molecules, 27(24), 8824. https://www.mdpi.com/1420-3049/27/24/8824.
- Toma, T. I., & Islam, A. (2023). Efficient in vitro Regeneration of Brassica napus L. Var. BARI Sorisha-18. Plant Tissue Culture and Biotechnology, 33(2), 107-114. https://www.baptcb.org/public/article/753_article.pdf.
- Xie, T., Chen, X., Guo, T., Rong, H., Chen, Z., Sun, Q., Batley, J., Jiang, J., & Wang, Y. (2020). Targeted knockout of BnTT2 homologues for yellow-seeded Brassica napus with reduced flavonoids and improved fatty acid composition. Journal of agricultural and food chemistry, 68(20), 5676-5690. https://pubs.acs.org/doi/abs/10.1021/acs.jafc.0c01126.
- Yang, Y., Zhu, K., Li, H., Han, S., Meng, Q., Khan, S. U., Fan, C., Xie, K., & Zhou, Y. (2018). Precise editing of CLAVATA genes in Brassica napus L. regulates multilocular silique development. Plant biotechnology journal, 16(7), 1322-1335. https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.12872.
- Yirmibeş, B., & Ülger, N. (2024). Botanical Characteristics, Nutritional Value, and Economic Importance of the Brassica Genus. Journal of Agriculture, Food and Ecology, 2(1), 9-14. https://dergipark.org.tr/en/download/article-file/4078475.
- Zhai, Y., Yu, K., Cai, S., Hu, L., Amoo, O., Xu, L., Yang, Y., Ma, B., Jiao, Y., Zhang, C., Khan, M.H.U., & Zhou, Y. (2020). Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnology Journal, 18(5), 1153-1168. https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.13281.
- Zheng, M., Zhang, L., Tang, M., Liu, J., Liu, H., Yang, H., Fan, S., Terzaghi, W., Wang, H., & Hua, W. (2020). Knockout of two Bna MAX 1 homologs by CRISPR/Cas9-targeted mutagenesis improves plant architecture and increases yield in rapeseed (Brassica napus L.). Plant biotechnology journal, 18(3), 644-654. https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.13228.
- Zhu, M., Monroe, J. G., Suhail, Y., Villiers, F., Mullen, J., Pater, D., Hauser, F., Jeon, B.W., Bader, J.S., Kwak, J.M. and Schroeder, J.I., & Assmann, S. M. (2016). Molecular and systems approaches towards drought-tolerant canola crops. New Phytologist, 210(4), 1169-1189. https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.13866.
Figure 1.
Overview of Tissue Culture and Regeneration Process in Canola (Brassica napus L.).
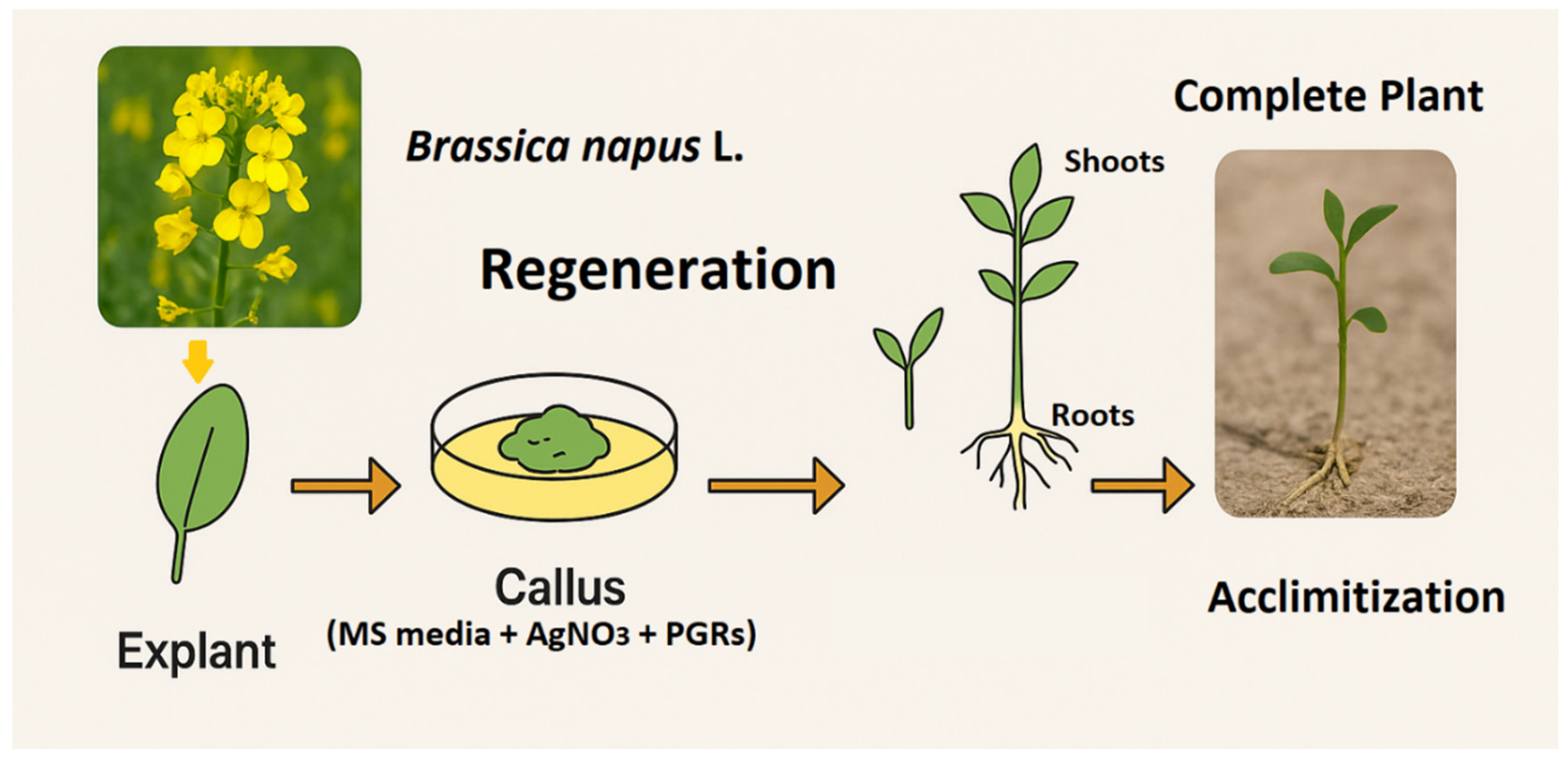
Figure 2.
Integrated Gene–Hormone (TFs) Regulatory Network Governing Shoot and Root Meristem Maintenance and Somatic Embryogenesis in Plants. Note: TFs=Transcription Factors (STM = SHOOT MERISTEMLESS, WUS = WUSCHEL, CUC = CUP-SHAPED COTYLEDON, BBM = BABY BOOM, LEC = LEAFY COTYLEDON, PLT = PLETHORA), ARFs = Auxin Response Factors, IAA = Indole-3-acetic acid (auxin), 2,4-D = 2,4-dichlorophenoxyacetic acid (synthetic auxin), GA = Gibberellin, SAM = Shoot Apical Meristem, RAM = Root Apical Meristem, SE = Somatic Embryogenesis, PGRs = Plant Growth Regulators, ↑=activates, ↓=suppress.
Figure 2.
Integrated Gene–Hormone (TFs) Regulatory Network Governing Shoot and Root Meristem Maintenance and Somatic Embryogenesis in Plants. Note: TFs=Transcription Factors (STM = SHOOT MERISTEMLESS, WUS = WUSCHEL, CUC = CUP-SHAPED COTYLEDON, BBM = BABY BOOM, LEC = LEAFY COTYLEDON, PLT = PLETHORA), ARFs = Auxin Response Factors, IAA = Indole-3-acetic acid (auxin), 2,4-D = 2,4-dichlorophenoxyacetic acid (synthetic auxin), GA = Gibberellin, SAM = Shoot Apical Meristem, RAM = Root Apical Meristem, SE = Somatic Embryogenesis, PGRs = Plant Growth Regulators, ↑=activates, ↓=suppress.
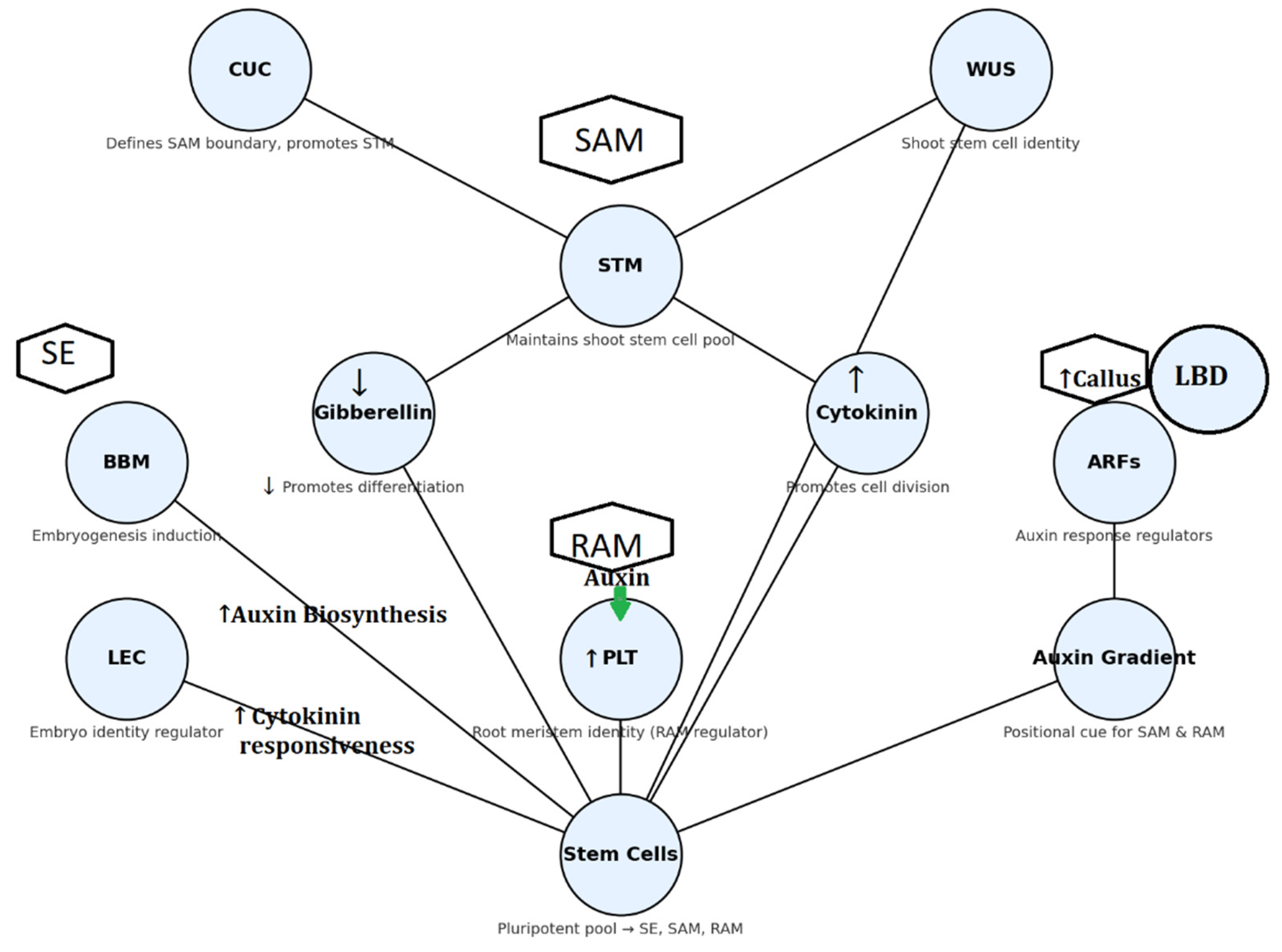
Figure 3.
Somatic Embryogenesis in Model and Medicinal Plants: A Comparative Framework.
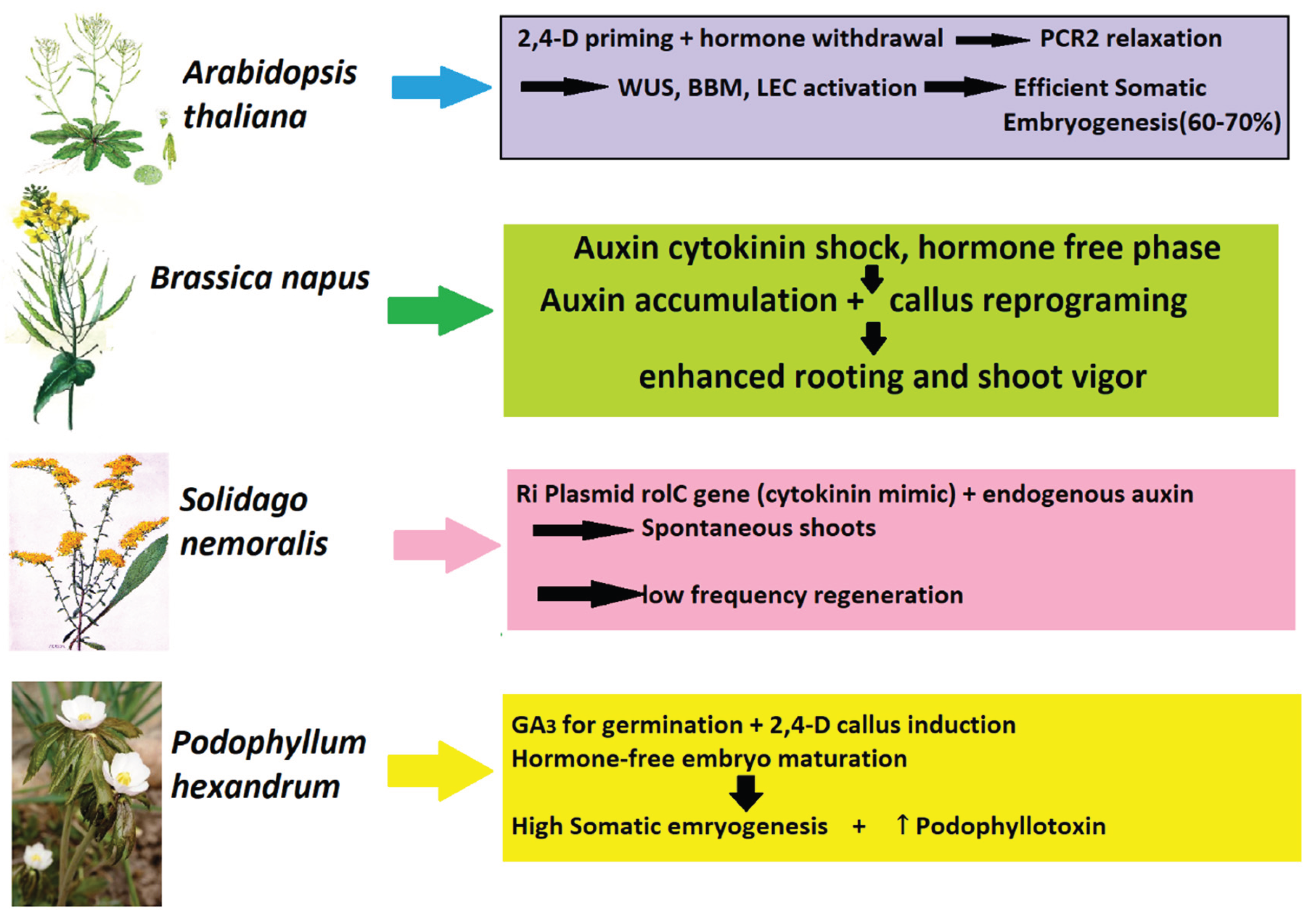

Table 1.
Explant types and their regeneration potential in Brassica napus.
| Explant Type | Shoots/explant | Limitations | Reported Regeneration Efficiency | References |
|---|---|---|---|---|
| Hypocotyl | Average 3.3 shoots/explant | Genotype-dependent | 0–50.6%, 82% | Farooq et al., 2019; Maheshwari et al., 2011; Dina et al., 2019; Ahmad et al., 2016 |
| Cotyledon | Average 9 shoots/explant | Genotype and protocol specific | 6–73%, 100% | Farooq et al., 2019; Dina et al., 2019; Lone et al., 2016; Biswas et al., 2017 |
| Petiole | Average 2.8 shoots/explant | Genotype and protocol specific | 4–79.3% | Farooq et al., 2019 |
| Roots | Average <2.35 shoots/explant | Very low shoot regeneration | 0–42.6% | Farooq et al., 2019 |
| Anther culture | 2.44shoots/callus | Genotype and protocol dependent | 40-60% | Sayem et al., 2010; Kumari et al., 2015 |
Table 2.
PGRs & combinations and their effects on canola regeneration.
| Hormone or Combination (MS Medium) | Explant used | Typical Effect on Regeneration | Remarks | References |
|---|---|---|---|---|
| 2,4-D (0.1 mg/L) | Hypocotyl | 1.4 g of callus obtained | 5mg/L silver nitrate extra ingredient | Naz et al., 2018 |
| BAP (2.5-3 mg/L) | Hypocotyl | 2.51 g callus (40-52.94%) | 5mg/l silver nitrate extra ingredient | Naz et al., 2018; Alam et al., 2014; Roh et al., 2012 |
| BAP (5.0 mg/l) and 2, 4-D (0.5 mg/l) | Cotyledon sections | 100% | Genotype dependent response | Lone et al., 2016 |
| 1.0 mg/l BAP and 0.1 mg/l NAA | Cotyledon and Hypocotyl | cotyledonary leaf with petiole (63.33%) and hypocotyl (43.33%) | Took 12-16 days for the induction of shoots | Toma and Islam, 2023 |
| TDZ (1.6 mg/L/ 8µM) | Cotyledon | 47% shoots regeneration | 8.5mg/L silver nitrate | Roh et al., 2012 |
| BAP (2 -5mg/L) + IAA (0.5 mg/L) | Hypocotyl | 84% shoots | 14.5 Number of shoots | Hussain et al.,2014 |
| 0.1 mg/1 NAA and 0 to 4 mg/1 kinetin | Cotyledon and Hypocotyl | Roots Cot (72%) Hyp (81%) Shoots l(20%) |
Calli were induced in 2,4-D and NAA | Anonymous |
Table 3.
Comparative Effects of PGRS-Free Shock and Internal Reprogramming on Plant Regeneration.
| Plant / Cultivar | Shock Duration / Treatment | Observed Effect | Underlying Mechanism | Reference |
|---|---|---|---|---|
| Arabidopsis thaliana (immature zygotic embryos) | 7 days on hormone-free MS after 2,4-D exposure | Efficient induction of somatic embryos (~60–70% ZEs); prolonged culture produced callus-like tissue and some shoots | 2,4-D primes dedifferentiation; removal allows IAA gradient establishment; PRC2 relaxation (in mutants or by transient depletion) removes epigenetic barrier, enabling embryo TF activation and reprogramming | Mozgová et al., 2017 |
| Brassica napus (Cyclone, Star, Westar explants) | Temporary 7-day “shock” on hormone-free MS after 0.5 mg/L 2,4-D + 0.5 mg/L BAP exposure | Enhanced rooting on 0.1 mg/L NAA + 2 mg/L BAP; improved shoot vigor | Hormone-free shock resets endogenous auxin/cytokinin balance; enhances auxin accumulation for rooting; mimics Arabidopsis withdrawal phase | Khan et al., 2010; Khan et al., 2024 |
| Solidago nemoralis (hairy roots, Agrobacterium rhizogenes–transformed, on 400 mg/L cefotaxime + sucrose) | 4–6 weeks | Low-frequency spontaneous shoot regeneration (2–3 shoots per hairy root); NAA further improved shoot regeneration | Endogenous auxin combined with internal reprogramming via rolC (cytokinin-mimetic effect); this internal hormone balance increased regeneration | Gunjan et al., 2013 |
| Podophyllum hexandrum (Milam ecotype seeds, cotyledon explants) | GA₃ (5 mg/L) for germination → 2,4-D (1.5 mg/L) in dark for callus induction → hormone-free MS for embryo maturation | Very high SE frequency (89.6% direct SE; 47.7 embryos per 50 mg callus); 79% mature embryos germinated; higher podophyllotoxin (1.8 mg/g DW) | GA₃ breaks dormancy & promotes germination; 2,4-D induces embryogenic callus; hormone-free phase allows embryo polarity; stress + dedifferentiation boost secondary metabolism (podophyllotoxin) and embryogenic competence | Rajesh et al., 2014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



