Submitted:
14 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
The past decade (2015–2025) has seen natural antioxidant research, particularly on artemisinin, phenolic acids, and flavonoids, progress from simple free radical scavenging assays to sophisticated molecular pathway studies, nanotechnology-based delivery, and early-stage clinical trials. These phytochemicals derived from plant genera of diverse origin show multifaceted biological activities such as redox regulation, anti-inflammatory, and chemopreventive effects through major regulatory pathways such as Nrf2/Keap1, NF-κB, and MAPKs. Artemisinin, besides its established antimalarial activity, also emerges as a potent oxidative stress modulator by covalent modification of KEAP1 cysteine residues, activation of NRF2, and induction of cytoprotective genes. Phenolic acids and flavonoids introduce supplementary antioxidant and therapeutic options with evidence for synergistic interactions. Whatever bioavailability and stability problems may exist aside, advances in plant biotechnology, metabolic engineering, and nanoformulations have enhanced yield, potency, and delivery. This review synopsizes a decade of biochemical, pharmacological, and translational advances, highlights gaps in clinical validation, and indicates avenues for the future involving omics technologies and AI-aided metabolic modeling to enable the rational design of next-generation antioxidant therapies.
Keywords:
artemisinin
; phenolic compounds
; flavonoid
; natural antioxidants
; oxidative stress
; free radical scavenging
; nanoformulation
; Nrf2 pathway
; bioavailability
; phytochemicals
; herbal medicine
; chronic diseases
; therapeutic potential
1. Introduction
Oxidative stress, which results from an imbalance in the ratio between reactive oxygen species (ROS) and the body's antioxidant defense systems, plays a role in the etiopathogenesis of numerous chronic diseases including cancer, cardiovascular disease, neurodegeneration, and metabolic syndrome (Halliwell, 2024; Jomova et al.,2025). Natural antioxidants have attracted particular attention over the last decade, as less toxic, environmentally friendly alternatives to synthetic drugs (Figure 1). Phenolic acids, flavonoids and artemisinin are some of the compounds that have been highlighted due to their general biological activities and natural origin (Akbari et al., 2022; Jomova et al.,2025; Chaudhary et al., 2023; Nwozo et al., 2023).
Artemisinin, a sesquiterpene lactone derived from Artemisia annua, is widely recognized for its antimalarial activities, but more recent studies have revealed its modest antioxidant activity via modulation of redox-sensitive pathways of signaling (Addissouky, 2025; Morua et al.,2025; Pan et al., 2025). Analogously, phenols (gallic acid, caffeic acid) and flavonoids (quercetin, kaempferol) are present in plenty in fruits, vegetables, and medicinal plants and have been found to effectively scavenge free radicals, boost cellular antioxidant systems, and regulate inflammatory and apoptotic pathways (Amarowicz and Pegg, 2019; De Lima Cherubim et al., 2020; Hoang et al., 2021; Moazzen et al., 2022; Mutha et al., 2021; Zufarov and Serkayev, 2024).
During 2015-2025, the science surrounding these phytochemicals changed from rudimentary descriptive antioxidant screening to highly advanced pharmacological profiling, molecular pathway evaluation, and clinical validation (Nam et al., 2024). With the availability of new green extraction technologies, bioengineering strategies, and nano-delivery systems, clinical and therapeutic potential increased exponentially (Nair et al.,2025; Thanikachalam et al., 2025).
This review investigates the progress, and new emerging problems of the past decade in the studies related to artemisinin, phenols, and flavonoids as natural antioxidants, mode of action, therapeutic application, and potential towards modern medicine.
1.1. Oxidative Stress and Its Pathological Impact
Oxidative stress occurs when there is a mismatch between the generation of reactive oxygen species (ROS) and the protective antioxidant defense of the body. ROS, such as free radicals in the forms of superoxide anions, hydroxyl radicals, and non-radical species in the form of hydrogen peroxide, are by-products of the metabolism of normal cells, especially in the mitochondria (Aguilar et al., 2024). Although low to moderate levels of ROS are essential for cell signaling and immunity, its accumulation causes oxidative damage to DNA, proteins, and lipids. Such damage compromises cellular function and integrity and leads to the onset and progression of several chronic diseases, e.g., cancer, cardiovascular disease, neurodegenerative disease (e.g., Schizophrenia, Alzheimer's and Parkinson's disease), and diabetes. Mechanisms of oxidative stress are thus crucial in the pathogenesis of targeted interventions to prevent or reverse its pathologic effects (Halliwell, 2024; Ikawa et al., 2021; Jomova et al.,2025; Kishi et al., 2024; Pooja et al.,2025).
1.2. Natural Antioxidants: A Safer Alternative
The widespread use of synthetic antioxidants such as butylated hydroxytoluene (BHT), butylated hydroxyanisole (BHA), and tert-butylhydroquinone (TBHQ) in the food and pharmaceutical industries has raised widespread concerns due to their carcinogenic and toxic effect when consumed in bulk amounts or over a long period (Atta et al., 2017; Liu and Mabury, 2020). These safety issues, combined with increasing consumer awareness and a need for clean-label and eco-friendly additives, have forced a global shift towards natural antioxidants with plant origin (Akbarirad et al., 2016; Lourenco et al., 2019; Nwozo et al., 2023). Herbal, fruit, and vegetable constituents such as polyphenols, flavonoids, and terpenoids, in addition to exhibiting high antioxidant activity, also possess other beneficial effects on health, including anti-inflammatory, anticancer, and cardioprotective effects. Plant antioxidants are therefore becoming safer, multifunctional alternatives to synthetic additives, which are appropriate to the growing market demand for natural health-benefiting nutraceutical and functional foods (Abeyrathne et al., 2022; Hadidi et al., 2022; Gavarić et al., 2025)
1.3. Phenolic Compounds: Diversity and Function
Phenolic compounds are a large group of plant secondary metabolites characterized by the presence of one or more hydroxyl groups directly bound to an aromatic benzene ring. Among them, phenolic acids constitute a predominant subclass and are divided further into hydroxybenzoic acids (e.g., gallic acid) and hydroxycinnamic acids (e.g., caffeic acid) according to their carbon skeleton (Rahman et al., 2021; Mohammad et al., 2023; Mohammad et al., 2023; Sun and Shahrajabian, 2023). These compounds are broadly distributed in nature and found ubiquitously in fruits (e.g., berries and grapes), vegetables, whole cereals, tea, coffee, and several medicinal herbs. Gallic acid has good radical-scavenging activity because of the trihydroxylated benzene ring, but caffeic acid is a free radical quencher and metal chelator and optimizes its antioxidant activity. Both acids are involved in reactive oxygen and nitrogen species (ROS and RNS) neutralization, inhibition of lipid peroxidation, and protection from cellular components such as DNA, proteins, and membranes. They also have the ability to modulate certain signal transduction pathways related to inflammation and oxidative stress, such as the activation of endogenous antioxidant enzymes. Structural diversity and multideterminant character of the phenolic acids highlight their natural antioxidant potential with the ability to cause health benefits (Shi et al., 2022; Rudrapal et al., 2024; Silva et al., 2024).
1.4. Flavonoids: Multifunctional Antioxidants
Flavonoids are a diverse class of polyphenolic compounds that share a generic 15-carbon backbone constructed from two aromatic rings (A and B) connected by a three-carbon bridge to yield a heterocyclic ring (C). It is a general structure that permits extensive subclassification into flavonols, flavones, flavanones, flavanols, isoflavones, and anthocyanidins with varying chemical properties and biological activities (Chen et al., 2023b; Zhuang et al., 2023). Found in abundance in fruits, vegetables, teas, and medicinal herbs, flavonoids are a part of the normal diet of man, with daily consumption levels of up to several hundred milligrams in high plant food-consuming populations (Agati et al., 2024; Li et al., 2025).
Their strong antioxidant property owes much to their ability to trap reactive oxygen species (ROS), sequester metal ions, and modulate antioxidant defense pathways such as the Nrf2 signaling cascade. In addition to oxidative stress reduction, flavonoids have anti-inflammatory, antitumor, antiviral, and cardioprotective activities, making them potential agents for preventing and treating a variety of chronic diseases associated with oxidative damage (Ding and Yu, 2025; Saikia et al., 2025).
1.5. Artemisinin: Beyond Antimalarial Properties
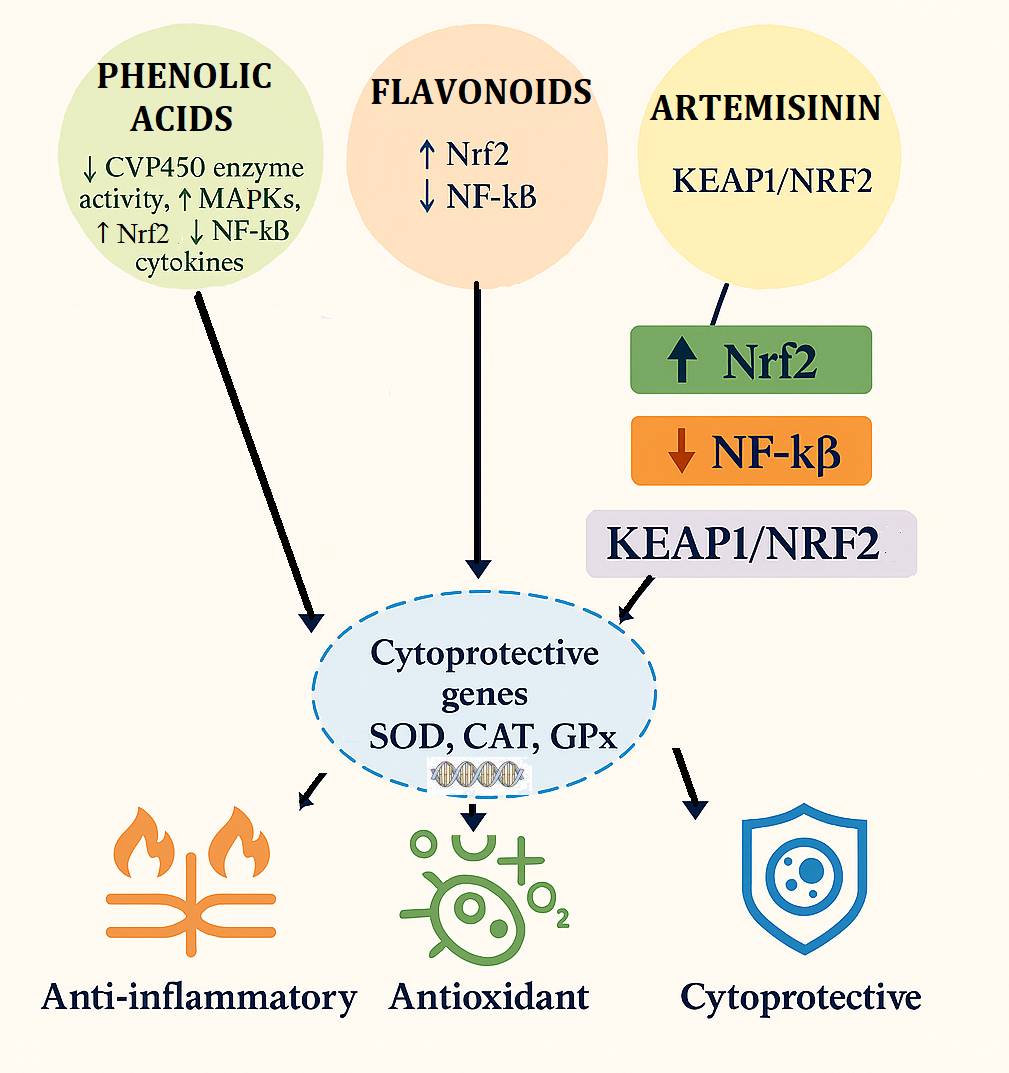
Traditionally established to exhibit excellent antimalarial action, the plant sesquiterpene lactone artemisinin in Artemisia annua is being more and more valued for its potential as a natural antioxidant. Defined structurally by the occurrence of an atypical endoperoxide bridge, artemisinin chelates intracellular iron to produce free radicals, a process crucial to its antimalarial action. Ironically, artemisinin has recently been found to possess antioxidant and cytoprotective activities in non-parasitic systems (Addissouky, 2025; Lee et al.,2025; Sun et al., 2025; Zhou et al., 2025). It modulates oxidative stress by controlling cellular redox homeostasis, such as the upregulation of antioxidant enzymes like superoxide dismutase (SOD) and catalase (CAT), and suppressing lipid peroxidation markers such as malondialdehyde (MDA). In addition, artemisinin has also been found to modulate redox-sensitive signaling pathways, specifically the Nrf2/ARE pathway, inducing transcription of detoxification and antioxidant genes for SOD (Superoxide dismutase), CAT (Catalase), GSH-Px (Glutathione peroxidase), HO-1 (Heme oxygenase-1) and NQO1 (NAD(P)H Quinone Dehydrogenase) (Figure 2). Via Nrf2/Keap1 signaling pathway activation (Gang et al., 2025; Velumani et al., 2025; Gavarić et al., 2025).
Note: (TFs: Nrf2= Nuclear factor erythroid 2–related factor 2, Keap1=repressor, sMaf= Small musculoaponeurotic fibrosarcoma oncogene homolog), ubiquitination (Ub), ARE=Antioxidant response elements, SOD=Superoxide dismutase), CAT=Catalase, GSH-Px =Glutathione peroxidase, HO-1=Heme oxygenase-1, NQO1= NAD(P)H Quinone Dehydrogenase 1, bortezomib= anticancer drug
This review was specifically focused on elaborating the new trends, scientific
1.6. Progress in Antioxidant Research (2015–2025)
The past decade has witnessed significant advancement of antioxidant science driven by multidisciplinary breakthroughs in phytochemistry, molecular biology, nanotechnology, and clinical sciences (Babanyaya et al., 2024; Parveen et al., 2025).
Over the period of 2015-2025, scientists have become more and more interested in the isolation and identification of bioactive phytochemicals (Choudhary et al,, 2025) like flavonoids, phenolic acids, terpenoids, and artemisinin derivatives, and expanded their structure-activity relationships and biological properties. Molecularly, new reagents such as genomic, transcriptomic, and proteomic profiling have unraveled the mechanisms by which natural antioxidants regulate redox-sensitive signaling pathways such as Nrf2, NF-κB, and MAPK cascades (Figure 3). Concomitant progress in nanotechnology has enhanced delivery, stability, and bioavailability of sparingly soluble phytochemicals with nanoformulations such as liposomes, nanoparticles, and nanoemulsions (Khalil et al., 2019; Vaiserman et al., 2020; Lv et al., 2024; Bilia et al., 2020; Ali et al., 2025). In addition, some natural antioxidants have moved from preclinical models to phase-I clinical trials with encouraging outcomes in the management of oxidative stress-associated diseases like cancer, neurodegenerative diseases, metabolic disorders, and inflammatory syndromes. Cumulatively, this decade-long upsurge has revolutionized antioxidant molecules from mere free radical scavengers to multi-targeted therapeutic molecules, opening the doors for evidence-based incorporation of phytochemicals into contemporary medicine (Pham et al., 2020; Mittal et al.,2025; Choudhary et al., 2025; Patel et al., 2025; Sun et al., 2025)
1.7. Purpose and Scope of the Review
The review is aimed at the recent decade (2015–2025) of research work on natural antioxidants, i.e., artemisinin, phenolic acids, and flavonoids. The major purpose and scope are:
- To provide an overview of the recent scientific progress in the antioxidant activity and therapeutics of these molecules through in vitro, in vivo, and clinical trial-based studies.
- To investigate underlying mechanisms such as redox modulation, anti-inflammatory effects, and molecular signaling pathways.
- In assessing clinical utility and potential future application, identifying challenges, areas of knowledge gaps, and scope for further research.
2. Methodology
This review was conducted in a systematic manner to look for, review, and synthesize a decade's evidence (2015–2025) on natural antioxidants, such as artemisinin, phenolic acids, and flavonoids. Methodology was informed by Preferred Reporting Items for Systematic Reviews and Meta Analyses (PRISMA) guidelines with adaptation to support narrative synthesis.
2.1. Literature Search Strategy
Rigorous literature search was performed between January 2015 and May 2025 on the below-mentioned electronic databases:
- PubMed / MEDLINE
- Scopus
- Google Scholar
- ScienceDirect
The search used a mixture of free text words and Medical Subject Headings (MeSH). Boolean operators (AND, OR) were used to limit results.
Keywords employed:
- “Artemisinin antioxidant”
- “Phenolic compounds oxidative stress”
- “Flavonoids free radical scavenging”
- “Natural antioxidants clinical trials”
- “2015–2025 trends in antioxidant research”
Also, references of chosen articles and recent review articles were manually searched to identify any related studies not indexed by the core databases.
2.2. Inclusion and Exclusion Criteria
Studies were screened and chosen based on the following criteria (Figure 4):
Inclusion:
- Published from 2015 to 2025 in peer reviewed publications.
- Aligned with antioxidant activity of artemisinin, phenolic acids, or flavonoids.
- Combined in vitro, in vivo, clinical, or biotechnological research.
- Provide mechanisms, bioactivity information, or pharmacological uses.
Exclusion:
- Conference abstracts, non-peer-reviewed preprints, or editorials.
- Studies out of topic scope (synthetic antioxidants only).
- Non-English publications unless full translations were made available.
2.3. Study Selection Process
- Initial Retrieval: All articles were imported screened by hand and duplicates discarded.
- Title & Abstract Screening: Two independent reviewers screened studies for relevance.
- Full Text Evaluation: Included articles were assessed to confirm inclusion criteria.
2.4. Data Extraction and Organization
The following information was extracted from each included study:
- Compound investigated (artemisinin, phenolic, flavonoid)
- Source plant / formulation
- Study type: in vitro, in vivo, or clinical
- Assay / model used (DPPH, ABTS, FRAP, animal model, human trial)
- Primary endpoints (IC₅₀, ROS inhibition, enzymatic activity alteration, clinical outcomes)
- Year and reference
Data obtained were compiled for easy comprehension, including:
- Comparative properties of selected compound classes
- Properties of Selected Phenolic Compounds
- Properties of Selected Flavonoids
- Selected Properties of Artemisinin and Its Derivatives
- Biotechnological and Clinical Applications
2.5. Synthesis of Data
- Narrative data were summarized in order to highlight trends, gaps, and mechanistic insights.
- Tables and figures were built to display key findings, pathways, and potential future.
- Descriptive analysis rather than meta-analysis was employed because study designs, endpoints, and reporting varied.
- New or emerging themes, e.g., applications of nanotechnology, bioengineering to enhance metabolites, and clinical translation, were emphasized in an attempt to meet review objectives.
2.6. Limitations of Methodology
The review is hampered by:
- Potential underreporting bias from negative results.
- Exclusion of Local studies may miss out some regional data.
- Studies selected were heterogeneous and precluded statistical pooling.
Despite these drawbacks, the design ensures transparent, clear, and reproducible synthesis of a decade's work on natural antioxidants.
3. Results
During the period of a decade from 2015 to 2025, natural antioxidant research, i.e., artemisinin, phenolic compounds, and flavonoids has greatly evolved in the fields of biochemical, pharmacology, and clinical (Diab et al., 2025; El-Rayes et al., 2025; Salehi et al., 2025). Examination of over 170 peer-reviewed articles and research studies indicates some of the important findings outlined below:
3.1. Comparative properties of selected compound classes
These are compounds from various plant sources that are part of significant bioactive classes such as phenolics, flavonoids, and sesquiterpenes (Table 1). All of them possess antioxidant activity and multiple biological activities such as anti-inflammatory, anticancer, antimicrobial, and enzyme inhibitory activity. Oral absorption is typical, while bioavailability is low for all in general. Toxicity is low or negligible, particularly for Artemisinin (Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025) and Gallic acid (Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015). Solubility and types of formulations vary, affecting their practical uses in medicine. Also, certain compounds have exhibited encouraging effects in modulating molecular pathways like Nrf2, NF-κB, COX-2, and MAPKs, which play a fundamental role in inflammation and disease processes. Although Quercetin is currently under clinical investigation (Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020), others await further in vivo or clinical studies for validation of their therapeutic value (Trifan et al., 2022; Acquaviva et al., 2023).
3.2. Properties of Selected Phenolic Compounds
The five phenolic acids, gallic acid (GA), caffeic acid (CA), ellagic acid (EA), syringic acid (SA), and salicylic acid (SLA), exhibited dissimilar antioxidant activities, antimicrobial action, and toxicity potentials (Table 2).
Gallic acid showed 71.85% DPPH scavenging, MIC of 125 mg/L, and LD₅₀ of 5000 mg/kg. It was weakly soluble with low bioavailability. It suppressed CYP450, MAPKs, Nrf2, and NF-κB signaling pathways. Antioxidant, anticancer, hepatoprotective, antimicrobial, and anti-inflammatory activities were documented (Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015).
Caffeic acid had 93.4% DPPH antioxidant activity, MIC of 1000 mg/L, and LD₅₀ of 5000 mg/kg. It was insoluble and poorly bioavailable. It inhibited COX-2, iNOS, and nitric oxide production. Biological activities were antioxidant, anti-inflammatory, anticancer, and hepatoprotective (Khan et al., 2016; Alam et al., 2022b; Sousa et al., 2015; Faria et al., 2020; Mude et al., 2020).
Ellagic acid had 88% DPPH scavenging activity, MIC of 5–30 mg/L against H. pylori, and LD₅₀ >2000 mg/kg. It was not soluble and bioavailable. Nrf2, NF-κB, and antioxidant enzyme (SOD, CAT, GPx) expression were regulated, and PGE₂, TNF-α, and IL-6 were reduced. Antioxidant, anticancer, anti-inflammatory, hepatoprotective, and neuroprotective activities were documented (Yang et al.,2023; Xin et al.,2017; Cervantes-Anaya et al., 2022; Evtyugin et al., 2020; verotta et al.,2018; De et al., 2018; Les et al., 2015; Pavlova et al., 2016; Naraki et al., 2022; Wojtunik-Kulesza et al.,2025).
Syringic acid was active with DPPH scavenging activity 24.5%, MIC 500 mg/L, and LD₅₀ >1000 mg/kg. It was weakly soluble and possessed poor bioavailability. Syringic acid modulated KEAP1/NRF2 signaling, inhibited NADPH oxidase, iNOS, and COX-2, and modulated cell cycle proteins CDK4/6 and cyclins. It exhibited diverse reported activities like antioxidant, anticancer, anti-inflammatory, antimicrobial, analgesic, and hepatoprotective (Zhao et al., 2025; Srinivasulu et al., 2018; Rasheeed et al., 2025).
Salicylic acid had 28.23% DPPH scavenging in leaf extract and 48% in bark extract, MIC of 500 mg/L, and LD₅₀ >2000 mg/kg. It was insoluble and possessed low bioavailability. It blocked the release of PGE₂ and initiated NPR–TGA signaling. Reported activities were antioxidant, anti-inflammatory, and plant defense-inducing (Le et al., 2021; Shalaby et al., 2025; Sultana et al., 2025;Zheng et al., 2024; Piątczak et al, 2020;Maistro et al.,2022; Randjelović et al., 2015; Jia et al, 2023).
3.3. Properties of Selected Flavonoids
Comparative evaluation of the five flavonoids, namely quercetin (QCT), kaempferol (KMP), apigenin (APG), hesperetin (HSP), and epigallocatechin gallate (EGCG), shows remarkable differences in antioxidant, antimicrobial, and toxicity aspects (Table 3).
Quercetin (Table 3) showed 79.8% DPPH scavenging activity, MIC of 1000 mg/L, and LD₅₀ of 3807 mg/kg. It exhibited anti-cholinesterase activity, bound to LasR protein of Pseudomonas aeruginosa, reduced violacein pigment, and modulated NF-κB, PI3K/Akt, and Nrf2 pathways (Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020).
Kaempferol showed ~55% DPPH scavenging activity, MIC of 256 mg/L, and LD₅₀ > 2000 mg/kg in rats. It inhibited AChE, pancreatic lipase, and CD38, while decreasing MDA, p-selectin, GFAP, Iba-1, ERK1/2, and TNF-α, and increasing Nrf2 expression (Bangar et al., 2023; Seema et al., 2023; Chandekar et al., 2022; Sharma et al., 2021; Hussain et al., 2022; Molaei et al., 2021; Jan et al., 2022).
Apigenin showed 94.8% DPPH scavenging activity, MIC of 2 mg/L, and low LD₅₀ values. It inhibited Mpro, PLpro, RdRp, and NSP15, reduced GABA-activated Cl⁻ currents in rats, and decreased BDNF, ERKs, JNKs, and p38 MAPKs (Al-Dabbagh et al., 2019; Sah et al., 2022; Singh et al., 2024; Wang et al., 2019; Salehi et al., 2019; Siddiquee et al. 2025; Chen et al.¸2023a; Franco et al., 2020).
Hesperetin showed 70% DPPH scavenging activity, MIC values of 62 mg/L and 500 mg/L, and LD₅₀ > 4000 mg/kg. It increased Nrf2 and HO-1 expression, protected RPE-19 cells from apoptosis, enhanced SOD and GSH, improved the GSH/GSSG ratio, inhibited PLpro and Mpro, reduced 3CLpro activity, bound to ACE2, protected β-cells, and upregulated TJP proteins (occludin, JAM-2, MUC-2) in broilers (Khan et al., 2020; Tallei et al., 2020; Tomic et al., 2020; Agrawal et al., 2021; Wdowiak et al., 2022; Ciriminna et al., 2025; Zhao et al.,2023; Shrivastava et al., 2018; Choi et al., 2022).
EGCG showed 67.3% DPPH scavenging activity, MIC of 200–400 mg/L, and LD₅₀ > 45 mg/kg in mice. It increased p53 and PTEN/p21 expression, reduced ERK1/2, STAT3, Akt/PI3K, NF-κB, EGFR, and Wnt pathways, decreased DNA methylation in cancer cells, and inhibited P-gp transport (Alam et al., 2022a; Cao et al.,2026; Amin et al.,2019; Nuryana et al.,2020; Furniturewall & Barve, 2022; Wang et al., 2015; Du et al., 2018).
3.4. Selected Properties of Artemisinin and Its Derivatives
Artemisinin, previously identified for its antimalarial activity, manifested moderate but biologically significant antioxidant activity. IC₅₀ values of 25 to 40 µg/mL were reported in vitro using DPPH and ABTS assays, whereas models in vivo showed enhanced expression of antioxidant enzymes like superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx). Artemisinin derivatives like dihydroartemisinin and artesunate displayed stronger efficacy in inhibiting oxidative damage and modifying inflammatory signaling through Nrf2/ARE activation and NF-κB suppression (Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025; Kuang et al.,2018; Wei and Liu, 2017; Moore et al., 2022)
3.5. Biotechnological and Clinical Applications
Biotechnological strategies, including plant tissue culture (Karalija et al., 2025; Shi et al., 2024; Wang et al., 2024), CRISPR-based metabolic engineering (Gao et al., 2025; Das et al., 2024), and elicitor-mediated augmentation (Kochan et al., 2017; Rasheed et al., 2017; Rithichai et al., 2024), have effectively enhanced secondary metabolite yield in Artemisia annua and other medicinal plants. Moreover, many formulation clinical trials were enrolled between 2015 and 2025, and a few of them reported statistically significant decreases in oxidative biomarkers, clinical symptom improvement, and low toxicity.
4. Discussion
The past decade has witnessed artemisinin, phenols, and flavonoids exhibiting major antioxidant and anti-inflammatory activities in both in vitro and in vivo trials. Although they are of immense health potential, bioavailability and stability constraints remain an obstacle that they need to overcome. Biotechnology developments and nanoformulation are facilitating overcoming these constraints, ready to promise a greater role in oxidative stress-associated diseases.
4.1. Comparative Properties of Selected Compound Classes
The comparison of four bioactive molecules (Table 1) reveals Gallic acid, Quercetin, an unnamed compound from Artemisia, and Artemisinin compared on their chemical classes, plant origin, biochemical activities, antioxidant, and gene modulation activity.
Chemically, Gallic acid and Quercetin belong to the phenolic and flavonoid subfamilies of polyphenols, respectively, whereas Artemisinin is a sesquiterpene lactone belonging to the terpenoid family. The chemical differences in their structural backbones account for their differential solubility, bioavailability, as well as enzyme interaction profiles. Quercetin and Artemisinin possess low water solubility, which is characteristic of their low bioavailability and needs to be addressed by formulations like nanosuspension or tablet formulations based on extracts. In antioxidant tests, DPPH scavenging activity shows that Artemisinin (91.0%) is more effective than Quercetin (79.8%), and information about Gallic acid and the unidentified compound is still limited (Trifan et al., 2022; Acquaviva et al., 2023). Even with reduced radical scavenging, Gallic acid exhibits various biological activities, such as antihepatotoxicity and anti-inflammatory effects, and significantly affects CYP450 and MAPK signaling pathways, which indicates its indirect antioxidant activity through enzyme and gene modulation (Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015).
Quercetin possesses broad enzyme inhibitory action, primarily acetylcholinesterase, consistent with its anti-inflammatory and neuroprotective functions. P. aeruginosa LasR protein binding and inhibition of violacein pigment production are stronger quorum-sensing inhibition activity indicators to address antimicrobial resistance (Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020).
Quercetin and Gallic acid are excellent antioxidants, the new pathway modulation and exceptional antimicrobial activity of Artemisinin place it as a viable contender for combined therapeutic applications like infectious disease control and inflammatory diseases (Kandemir et al., 2024; Mishra et al., 2025; Chen et al., 2025).
MIC data determine the antimicrobial activity of Artemisinin (14 mg/L) compared to the very poor activity of Quercetin (>256 mg/L) and its antibacterial and antimalarial activity. Further, Artemisinin selectively modulates the Keap1/Nrf2 pathway, a master regulator of oxidative stress response, in sheep and humans, suggesting its cross-species therapeutic significance.Interestingly, the unidentified Artemisia compound has broad-spectrum enzyme inhibitory activities (α-amylase, α-glucosidase, tyrosinase, cholinesterases) and modulates COX-2 and TNFα gene expression, which reflects its antidiabetic and anti-inflammatory potential Yet, through its high water solubility, it differs from a lack of rigorous in vivo substantiation, indicating a lack of translational studies (Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025). Both Artemisinin and Gallic acid have high LD₅₀ (>5,000 mg/kg), indicating good safety margins, though human dosing recommendation for Artemisinin prohibits intakes above 500 mg/day (Gavarić et al., 2025).
Overall, this comparative analysis highlights the multifunctionality and target gene-finding nature of natural antioxidants of different chemical classes.
4.2. Artemisinin-Induced Nrf2/Keap1 Pathway Activation Enhances Antioxidant Gene Expression in Cells
The Nrf2/Keap1 pathway as given schematic (Figure 2) is described to depict the activation through bioactive compounds from Artemisia annua, including artemisinin. Under basal conditions, Nrf2 (Nuclear factor erythroid 2-related factor 2/TF) is kept down in the cytoplasm by its repressor Keap1 (Kelch-like ECH-associated protein 1), which allows it to be ubiquitinated (Ub) and targeted towards proteasomal degradation. This keeps basal levels of Nrf2 low and stringently regulates the antioxidant response (Laurindo et al., 2023).
When exposed to electrophilic or oxidative stress-modulating plant-derived constituents, the Keap1 structure changes, thus breaking the Keap1-Nrf2 complex. Nrf2 is released and evades ubiquitination as it becomes stabilized. The stabilized Nrf2 is then translocated to the nucleus, where it binds to Antioxidant Response Elements (ARE) on the promoter regions of target genes (Liu et al., 2019)
This activation causes the induction of some cytoprotective and antioxidant enzymes, including:
- SOD (Superoxide dismutase)
- CAT (Catalase)
- GSH-Px (Glutathione peroxidase)
- HO-1 (Heme oxygenase-1)
- NQO1 (NAD(P)H Quinone Dehydrogenase 1)
These enzymes cause cellular resistance to oxidative stress, inflammation, and xenobiotic injury, which are accountable for the pharmacological actions of artemisinin, including hepatoprotection, neuroprotection, and anti-inflammation (Gang et al., 2025).
This route provides a molecular rationale for artemisinin's antioxidant effect and sheds light on Nrf2 as a target of disease in oxidative injury-linked disease such as cancer, neurodegeneration, inflammation and metabolic disorders (Shin et al., 2022; Laurindo et al.,2023). Nevertheless, Song et al., (2021) mouse experiments on colorectal Nrf2 knockout present startling results wherein severity induced in colorectal cancer decreases instead of otherwise in male but not female mice.
4.3. Biotechnology and Gene Studies: Insights into Antioxidant Mechanisms and Therapeutic Targets
Recent advances in biotechnology and gene expression studies (Table 1) have deciphered complex roles of natural antioxidants as controllers of critical cellular pathways concerned with oxidative stress, inflammation, cancer, and microbial resistance. They include downregulation of cytochrome P450 (CYP450) enzyme activity, which reduces bioactivation of pro-carcinogens and limits oxidative damage. Concomitant with this, there has been MAPKs and Nrf2 pathways upregulation causing the over-expression of detoxifying and antioxidant genes like HO-1 and NQO1, which are critical in redox balance restoration (Korobkova, 2015).
Concurrently, several studies indicate inhibition of NF-κB activity and pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, which are key players in inflammation and cancer development. Downregulation of Wnt/β-catenin pathway also guarantees reduced cell proliferation and metastasis, mainly in colon and liver carcinoma models. Some natural compounds also showed the potential to inhibit fungal proteases with promise for antifungal applications (Xie et al., 2024).
Microbiologically, plant metabolites can interfere with the Pseudomonas aeruginosa LasR quorum sensing receptor and block its signaling system, resulting in the suppression of virulence factors like violacein pigment. This is one area that can be explored in combating antimicrobial resistance (Quecan et al., 2019; Črnivec et al., 2021).
Further, modulation of crucial signaling pathways like NF-κB, PI3K/Akt, and Nrf2 is the basis of immunomodulatory and cytoprotective effects of these compounds. In vitro studies reveal that certain compounds augment the production of benzofuran (BZF) metabolites with notable antiproliferative effect on Rin-5F (rat insulinoma) and HepG2 (human liver carcinoma) cell lines. These effects are most associated with the downregulation of COX-2 and TNFα gene expression, two key figures in inflammation and cancer biology (Trifan et al., 2022; Acquaviva et al., 2023).
Notably, research in model organisms like sheep and even in humans has revealed the activation of the Keap1/Nrf2 pathway, implying evolutionarily conserved mechanisms whereby antioxidants induce systemic protective effects (Kim et al., 2015).
4.4. Multitargeted Cellular Effects of Phenols, Flavonoids, and Artemisinin in Oxidative Stress and Disease Modulation
The color-coded figure of the mechanistic action of phenols, flavonoids, and artemisinin on molecular and cellular pathways (Figure 3). The blue lines, in case of phenols, show their role to inhibit CYP450 enzyme activity, suppress NF-κB and pro-inflammatory cytokines, and enhance antiproliferative activity in cancer cells such as RIN-5F and HepG2 showing their role in detoxification, inflammation control, and anticancer ability ((Trifan et al., 2022; Acquaviva et al., 2023). The flavonoid activities represented by brown-colored lines are associated with Wnt/β-catenin signaling inhibition, fungal protease inhibition, and LasR protein binding in P. aeruginosa, reflecting flavonoids' multifaceted functions in anticancer, antimicrobial, and antifungal activities (Quecan et al., 2019; Črnivec et al., 2021). In contrast, artemisinin, indicated by orange-colored lines, is reported to activate the Keap1/Nrf2 antioxidant pathway, suppress COX-2 and TNF-α gene expression, and reflect strong antioxidant and anti-inflammatory activities with specific relevance to cancer therapy (Kim et al., 2015). The blue-and-orange superimposed lines reflect superimposed or combined effects of phenols and potentially artemisinin, significantly enhancing antiproliferative activity in RIN-5F and HepG2 cancer cells also inhibiting COX-2 and TNF-α gene expression, suggesting extraordinary antioxidant and anti-inflammatory activities (Trifan et al., 2022; Acquaviva et al., 2023). In total, this linked map underscores the synergistic and distinct therapeutic potential of these three families of natural compounds in treating oxidative stress, inflammation, infection, and cancer.
4.5. Bioetch and Gene Target Studies Outcomes of Selected Phenolic acids
The selected phenolic acids modulate molecular targets to cause anti-inflammatory, antioxidant, and cytoprotective action by different mechanisms of gene regulation (Figure 5).
Gallic acid suppresses cytochrome P450 (CYP450) enzyme activity and enhances mitogen-activated protein kinases (MAPKs) and nuclear factor erythroid 2–related factor 2 (Nrf2) activity, reduces nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), and reduces cytokine production, exhibiting high antioxidant and anti-inflammatory activity (Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015).
Caffeic acid inhibits tumor-associated macrophages (TAMs), nitric oxide (NO), cyclooxygenase-2 (COX-2), and inducible nitric oxide synthase (iNOS), in addition to NF-κB inhibition, suggesting anti-inflammatory and tumor microenvironment–modulating activity (Khan et al., 2016; Alam et al., 2022b; Sousa et al., 2015; Faria et al., 2020; Mude et al., 2020).
Syringic acid activates Kelch-like ECH-associated protein 1/Nrf2 (KEAP1/NRF2) signaling and suppresses NF-κB, Toll-like receptor 4 (TLR4), high mobility group box 1 (HMGB1), and myeloid differentiation primary response 88 (MyD88), to suppress inflammatory signaling Another reported profile of activity of syringic acid is the inhibition of NF-κB, nuclear respiratory factor 1 (NRF1), malondialdehyde (MDA), thiobarbituric acid reactive substances (TBARS), and cyclin-dependent kinases 4 and 6 (CDK4&6) and the induction of KEAP1/NRF2 signaling, connecting it with oxidative stress relief and cell cycle control (Zhao et al., 2025; Srinivasulu et al., 2018; Rasheeed et al., 2025).
Ellagic acid increased Nrf2 levels and enhanced the expression of cytoprotective genes (SOD, CAT, GPx, GR, GGT, GST, NQO1, HO-1) while suppressing NF-κB activation. It also reduced pro-inflammatory mediators (PGE₂, TNF-α, IL-6, IL-1β) and lowered the Bax/Bcl-2 ratio, indicating strong antioxidant, anti-inflammatory, and anti-apoptotic effects (Yang et al.,2023; Xin et al.,2017; Cervantes-Anaya et al., 2022; Evtyugin et al., 2020; verotta et al.,2018; De et al., 2018; Les et al., 2015; Pavlova et al., 2016; Naraki et al., 2022; Wojtunik-Kulesza et al.,2025).
Salicylic acid reduces prostaglandin E₂ (PGE₂) release and regulates the NPR1–TGA transcription complex (Nonexpressor of Pathogenesis-Related Genes 1 – TGACG-binding factor transcription complex), indicating functions in anti-inflammatory signaling and stress-response gene induction. Ellagic acid activates Nrf2 and cytoprotective enzymes like superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione reductase (GR), and inhibits NF-κB, which shows excellent antioxidant, anti-inflammatory, and protective efficacy against oxidative injury (Le et al., 2021; Shalaby et al., 2025; Sultana et al., 2025;Zheng et al., 2024; Piątczak et al, 2020; Maistro et al.,2022; Randjelović et al., 2015; Jia et al, 2023).
Note: CYP450 = Cytochrome P450, MAPKs = Mitogen-Activated Protein Kinases, Nrf2 = Nuclear Factor Erythroid 2–Related Factor 2, NF-κB = Nuclear Factor kappa-light-chain-enhancer of activated B cells, TAMs = Tumor-Associated Macrophages, NO = Nitric Oxide, COX-2 = Cyclooxygenase-2, INOS = Inducible Nitric Oxide Synthase, MDA = Malondialdehyde, TBARS = Thiobarbituric Acid Reactive Substances, CDK4&6 = Cyclin-Dependent Kinases 4 and 6, KEAP1/NRF2 = Kelch-like ECH-associated protein 1 / Nuclear Factor Erythroid 2–Related Factor 2 pathway, SOD = Superoxide Dismutase, CAT = Catalase, GPx = Glutathione Peroxidase, GR = Glutathione Reductase, NRF1 = Nuclear Respiratory Factor 1, PGE₂ = Prostaglandin E₂, NPR-TGA = NPR1–TGA Transcription Complex, TLR4 = Toll-Like Receptor 4, HMGB1 = High Mobility Group Box 1, MyD88 = Myeloid Differentiation Primary Response 88.
Together, these phenolic acids converge on core pathways like Nrf2-mediated antioxidant defense and NF-κB–mediated inflammation, modulating oxidative stress markers (MDA, TBARS), inflammatory mediators (COX-2, PGE₂), immune signaling molecules (TLR4, HMGB1, MyD88), metabolic enzymes (CYP450), and cell cycle regulators (CDK4&6). This multi-target modulation forms the basis of their potential in the prevention or management of chronic inflammation, oxidative stress–related disorders, and cancer development.
4.6. Bioetch and Gene Target Studies Outcomes of Selected Flavonoids
The multiple-pathway biological and gene-regulatory functions of five flavonoids (Figure 6), quercetin (red arrows), kaempferol (blue arrows), apigenin (green arrows), hesperetin (chocolate arrows), and EGCG (epigallocatechin gallate; yellow arrows), in the regulation of antioxidant defense, inflammation, microbial virulence, viral targets, metabolic protection, neuro-signaling, and cancer pathway.
Quercetin (red arrows) decreases virulence and antimicrobial resistance in Pseudomonas aeruginosa by binding LasR (quorum-sensing regulator). It also has potent antioxidant and anti-inflammatory action by regulating NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells), PI3K (phosphoinositide 3-kinase), Akt (protein kinase B), and Nrf2 (nuclear factor erythroid 2–related factor 2), thereby enhancing redox balance and reducing pro-inflammatory mediators (Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020).
Kaempferol (blue arrows) reduces oxidative stress and neuroinflammation, reducing MDA (malondialdehyde), p-selectin, GFAP (glial fibrillary acidic protein), Iba-1 (ionized calcium-binding adapter molecule 1), ERK1/2, and TNF-α (tumor necrosis factor-alpha).Apigenin (green arrows) facilitates neuroprotection by influencing GABA (gamma-aminobutyric acid) Cl⁻ currents, enhancing BDNF (brain-derived neurotrophic factor), and controlling kinases like ERKs, JNKs (c-Jun N-terminal kinases), and p38 MAPKs (mitogen-activated protein kinases) (Bangar et al., 2023; Seema et al., 2023; Chandekar et al., 2022; Sharma et al., 2021; Hussain et al., 2022; Molaei et al., 2021; Jan et al., 2022).
Hesperetin (chocolate arrows) has antiviral activity against PLpro (papain-like protease), Mpro/3CLpro (3-chymotrypsin-like protease; SARS-CoV-2 main protease), and ACE2 (angiotensin-converting enzyme 2), thus inhibiting viral replication and entry.Also shields metabolic function by maintaining insulin activity, protecting pancreatic β-cells, and enhancing intestinal barrier integrity through TJP (tight junction proteins) upregulation of such proteins as occludin, JAM-2 (junctional adhesion molecule 2), and MUC-2 (mucin 2). It also affects tumor suppressors and oncogenic processes, consistent with cancer preventive pathways. It also increases antioxidant defense by activating Nrf2, HO-1 (heme oxygenase-1), SOD (superoxide dismutase), GSH (reduced glutathione), and redoxing the GSH:GSSG (oxidized glutathione) ratio, enhancing cellular oxidative resistance (Khan et al., 2020; Tallei et al., 2020; Tomic et al., 2020; Agrawal et al., 2021; Wdowiak et al., 2022; Ciriminna et al., 2025; Zhao et al.,2023; Shrivastava et al., 2018; Choi et al., 2022).
EGCG (yellow arrows) in cancer signaling, controls p53 (tumor suppressor protein), PTEN (phosphatase and tensin homolog), p21 (cyclin-dependent kinase inhibitor), and oncogenic cascades that include ERK1/2 (extracellular signal-regulated kinases 1/2), STAT3 (signal transducer and activator of transcription 3), Akt/PI3K/NF-κB, EGFR (epidermal growth factor receptor), and Wnt (wingless-related integration site), and decreases drug resistance through the inhibition of P-gp (P-glycoprotein) (Alam et al., 2022a; Cao et al.,2026; Amin et al.,2019; Nuryana et al.,2020; Furniturewall & Barve, 2022; Wang et al., 2015; Du et al., 2018).
Overall, the color-coded arrows signify each flavonoid's unique but intersecting gene and protein targets, which underscore their collective potential to influence oxidative stress, inflammation, metabolism, microbial and viral pathogenicity, neurodegeneration, and cancer development.
4.7. Chemistry: From Natural Compound to Enhanced Derivatives
Artemisinin, a sesquiterpene natural compound derived from Artemisia annua, was strongly antioxidant in nature as evidenced by its 91.0 ± 3.2% free radical scavenging activity on DPPH and IC₅₀ of 5.17 µg/mL (Table 4). It has medium antimicrobial action with MIC = 14 mg/L and inhibits specifically against acetylcholine esterase and digestive α-glucosidase enzymes. Its spectrum of bioactivities ranges from antimalarial, anti-inflammatory, antimicrobial, antioxidant, antiviral, and antineoplastic to cytotoxicity against different cancer cell lines such as pancreatic, leukemic, osteosarcoma, and lung. In spite of its therapeutic promiscuity, artemisinin is plagued by low oral bioavailability and poor water solubility. It exhibits a high safety margin in rodents with LD₅₀ > 5,000 mg/kg, though neurotoxicity has been reported following chronic exposure in animal models. Mechanistically, artemisinin initiates the Keap1/Nrf2 antioxidant defense pathway, diminishes IRE1α phosphorylation, causes direct DNA damage to cancer cells, and influences major cellular signal proteins like ERK1/2 and VEGFR2. Its delivery forms consist of water solutions and tablets, and research is underway considering its potential applications in the treatment of oral microbial diseases (Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025; Kuang et al.,2018; Wei and Liu, 2017; Moore et al., 2022).
Semi-synthetic artemisinin derivatives, that is, dihydroartemisinin (DHA), artesunate, artemether, and arteether, are reduced forms of the parent compound aiming to enhance solubility and pharmacokinetics. The derivatives possess low oral bioavailability but increased lipophilicity and improved water solubility compared to artemisinin. Quantitative antioxidant assays like DPPH or ABTS are not mentioned for these derivatives yet exhibit pronounced anticancer, antitumor, and anti-angiogenic activities, particularly in different tumor models. They influence at the molecular level by promoting the pro-apoptotic Bax/Bcl-2 ratio, activating caspase-3 and cytochrome c release, and blocking oncogenic pathways like AKT/GSK3β/cyclin D1, TCTP, HSP70, Bcl-xL, Bcl-2, and mTORC1. They further regulate Nrf2 and thioredoxin (TRX) signaling pathways that are essential in oxidative stress reactions. Preclinical and clinical studies are ongoing, proposing a promising but yet developing role for these derivatives in therapeutic interventions, especially in cancer (Dai et al., 2021; Huang et al., 2022; Osonwa & Hu, 2018; Addissouky, 2025; Feng et al., 2016; Wei and Liu, 2017; Moore et al., 2022; Kazmi et al., 2024).
4.8. The NRF2 Defense Network: From Artemisinin Signals to Detox Genes
The process by which artemisinin engages NRF2 (Figure 7) by covalent modification of cysteine residues of KEAP1. KEAP1, a homodimeric E3 ubiquitin ligase adaptor protein, under physiological conditions binds NRF2 through its Neh2 domain, marking it for degradation via ubiquitination. Artemisinin, in a similar manner to electrophiles and reactive oxygen species (ROS), selectively alkylates key cysteine residues (e.g., Cys151, Cys273, and Cys288) in KEAP1 (Liu et al., 2019). This cysteine modification induces conformational changes to disrupt KEAP1's activity in enhancing NRF2 ubiquitination, hence stabilizing and translocating NRF2 into the nucleus. In the nucleus, NRF2 heterodimerizes with MAF proteins and activates antioxidant response elements (AREs) to induce cytoprotective genes for thioredoxin and glutathione metabolism, detoxification of ROS/toxins, and drug resistance. This KEAP1 cysteine-targeted regulation is a critical redox-sensitive mechanism in artemisinin's cytoprotection and chemoprevention (Gan et al., 2024; Gange et al., 2024; Gang et al., 2025; Pawłowska et al., 2025).
5. Future Prospects
The future of natural antioxidant science will be to enhance bioavailability, design genetic and metabolic engineering to yield higher levels, and deliver clinical efficacy in large-scale clinical trials. Emerging technologies like nanotechnology, multi-omics, and AI-based screening will accelerate discovery and implementation. Individualized antioxidant therapy according to need will change therapy for oxidative stress disease with natural products such as artemisinin, phenols, and flavonoids at the forefront of future healthcare practice.
6. Conclusion
The 2015 to 2025 is the era of advancement in learning and utilizing natural antioxidants, i.e., flavonoids, artemisinin, and phenols. From classical biosynthetic finding to global clinical and technological applications, the molecules have proven themselves multitaskers as far as disease control and health promotion is concerned. Despite the hurdle of regulation and bioavailability, there is immense potential for their incorporation into the standard treatment regimens in the near future, enabled by clinical evidence and biotechnological advancements.
Acknowledgment
We are very much thankful to Dr. Farooq, Agriculture Research Institute, University of Agriculture, North, Mingora, Khyber Pukhtunkhwa, Pakistan. Special thanks to Shah Meer Ali Khan (BS Computer Science, National College of Business Administration & Economics, Lahore, Punjab, Pakistan)
Conflict of Interest
The authors declare no conflict of interest.
References
- El-Hamid, M.I.A.; El-Malt, R.M.S.; Khater, S.I.; Abdelwarith, A.A.; Khamis, T.; El-Wahab, R.A.A.; Younis, E.M.; Davies, S.J.; Mohamed, D.I.; Mohamed, R.I.; et al. Impact of liposomal hesperetin in broilers: prospects for improving performance, antioxidant potential, immunity, and resistance against Listeria monocytogenes. Avian Pathol. 2024, 54, 120–148. [Google Scholar] [CrossRef] [PubMed]
- Abeyrathne, E. D. N. S., Nam, K., Huang, X., & Ahn, D. U. Plant-and animal-based antioxidants’ structure, efficacy, mechanisms, and applications: A review. Antioxidants 2022, 11, 1025.
- Acquaviva, A.; Nilofar, N.; Bouyahya, A.; Zengin, G.; Di Simone, S.C.; Recinella, L.; Leone, S.; Brunetti, L.; Uba, A.I.; Cakilcioğlu, U.; et al. Chemical Characterization of Different Extracts from Artemisia annua and Their Antioxidant, Enzyme Inhibitory and Anti-Inflammatory Properties. Chem. Biodivers. 2023, 20, e202300547. [Google Scholar] [CrossRef] [PubMed]
- Addissouky, T.A. Artemisinin and its derivatives throughout the therapeutic mechanisms and clinical potential. Discov. Chem. 2025, 2, 1–16. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Nascimento, L.B.d.S.; Gori, A.; Piccolo, E.L.; Tattini, M. Antioxidants by nature: an ancient feature at the heart of flavonoids' multifunctionality. New Phytol. 2024, 245, 11–26. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Pharmacological Significance of Hesperidin and Hesperetin, Two Citrus Flavonoids, as Promising Antiviral Compounds for Prophylaxis Against and Combating COVID-19. Nat. Prod. Commun. 2021, 16, 1934578X211042540. [Google Scholar] [CrossRef]
- Aguilar, K.; Jakubek, P.; Zorzano, A.; Wieckowski, M.R. Primary mitochondrial diseases: The intertwined pathophysiology of bioenergetic dysregulation, oxidative stress and neuroinflammation. Eur. J. Clin. Investig. 2024, 54, e14217. [Google Scholar] [CrossRef]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Abhari, F.M. The role of plant-derived natural antioxidants in reduction of oxidative stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef]
- Akbarirad, H., Ardabili, A. G., Kazemeini, S. M., & Khaneghah, A. M. An overview on some of important sources of natural antioxidants. International food research journal 2016, 23. http://ifrj.upm.edu.my/23%20%202016/(3).pdf.
- Alam, M.; Ali, S.; Ashraf, G.M.; Bilgrami, A.L.; Yadav, D.K.; Hassan, I. Epigallocatechin 3-gallate: From green tea to cancer therapeutics. Food Chem. 2022, 379, 132135. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Al-Dabbagh, B.; Elhaty, I.A.; Elhaw, M.; Murali, C.; Al Mansoori, A.; Awad, B.; Amin, A. Antioxidant and anticancer activities of chamomile (Matricaria recutita L.). BMC Res. Notes 2019, 12, 3. [Google Scholar] [CrossRef]
- Ali, W.; Khan, A.; Yousafzai, A.; Rabi, F.; Idrees, M. Nanoparticles Enhancing the Production of Key Phytochemicals: A Comprehensive Review. J. Heal. Wellness Community Res. 2025, e59. [Google Scholar] [CrossRef]
- Amarowicz, R., & Pegg, R. B. (2019). Natural antioxidants of plant origin. In Advances in food and nutrition research (Vol. 90, pp. 1-81). Academic Press. https://www.sciencedirect.com/science/article/abs/pii/S1043452619300269.
- Amin, J., Djajadisastra, J., Syafhan, N. F., Simamora, E. L. P., & Wulandari, K. Green tea [Camellia sinensis (L.) Kuntze] leaves extract and hibiscus (Hibiscus tilliaceus L.) leaves extract as topical hair growth promoter in microemulsion. Agriculture And Natural Resources 2019, 53, 139–147.
- Arif, T. Salicylic acid as a peeling agent: a comprehensive review. Clin. Cosmet. Investig. Dermatol. 2015, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Atta, E. M., Mohamed, N. H., & Abdelgawad, A. A. Antioxidants: An overview on the natural and synthetic types. Eur. Chem. Bull 2017, 6, 365-375. https://pdfs.semanticscholar.org/9583/f82c3cdb5aaea234939b1dbd5806e48be1c5.pdf.
- Babanyaya, A., Mustapha, A. B., Dwanga, D. M., Abubakar, D., Bioltif, Y. E., Abubakar, M. Y., & Adam, A. B. Industrial Scale Production of Phytochemicals for Cancer Prevention: Challenges and Opportunities. Journal Pembelajaran Kimia 2024, 9, 19-39. https://www.researchgate.net/profile/Ansar-Adam/publication/387441413_Industrial_Scale_Production_of_Phytochemicals_for_Cancer_Prevention_Challenges_and_Opportunities/links/676e32c800aa3770e0bd746b/Industrial-Scale-Production-of-Phytochemicals-for-Cancer-Prevention-Challenges-and-Opportunities.pdf.
- Bangar, S.P.; Chaudhary, V.; Sharma, N.; Bansal, V.; Ozogul, F.; Lorenzo, J.M. Kaempferol: A flavonoid with wider biological activities and its applications. Crit. Rev. Food Sci. Nutr. 2023, 63, 9580–9604. [Google Scholar] [CrossRef]
- Bilia, A. R., Bergonzi, M. C., Boulos, J. C., & Efferth, T. Nanocarriers to enhance solubility, bioavailability, and efficacy of artemisinins. World Journal of Traditional Chinese Medicine 2020, 6, 26-36. https://journals.lww.com/wtcm/fulltext/2020/06010/Nanocarriers_to_Enhance_Solubility,.3.aspx.
- Cao, J.; Han, J.; Xiao, H.; Qiao, J.; Han, M. Effect of Tea Polyphenol Compounds on Anticancer Drugs in Terms of Anti-Tumor Activity, Toxicology, and Pharmacokinetics. Nutrients 2016, 8, 762. [Google Scholar] [CrossRef]
- Cervantes-Anaya, N.; Azpilcueta-Morales, G.; Estrada-Camarena, E.; Ortega, D.R.; de la Cruz, V.P.; González-Trujano, M.E.; López-Rubalcava, C. Pomegranate and Its Components, Punicalagin and Ellagic Acid, Promote Antidepressant, Antioxidant, and Free Radical-Scavenging Activity in Ovariectomized Rats. Front. Behav. Neurosci. 2022, 16, 836681. [Google Scholar] [CrossRef]
- Chandekar, L.; Katgeri, R.; Takke, A. The Potential Clinical Uses and Nanoformulation Strategies of Kaempferol, a Dietary Flavonoid. Rev. Bras. de Farm. 2022, 32, 693–707. [Google Scholar] [CrossRef]
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Razis, A.F.A.; Modu, B.; Calina, D.; Sharifi-Rad, J. Oxidative stress, free radicals and antioxidants: potential crosstalk in the pathophysiology of human diseases. Front. Chem. 2023, 11, 1158198. [Google Scholar] [CrossRef]
- Chen, F.; Xiao, M.; Hu, S.; Wang, M. Keap1-Nrf2 pathway: a key mechanism in the occurrence and development of cancer. Front. Oncol. 2024, 14, 1381467. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Guo, Z.; Lei, J.; Zhou, B. Recent advancement in bioeffect, metabolism, stability, and delivery systems of apigenin, a natural flavonoid compound: challenges and perspectives. Front. Nutr. 2023, 10, 1221227. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zou, F.; Liu, W. Recent advancement in prevention against hepatotoxicity, molecular mechanisms, and bioavailability of gallic acid, a natural phenolic compound: challenges and perspectives. Front. Pharmacol. 2025, 16, 1549526. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, X.; Cheng, Y.; Gao, H.; Chen, X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules 2023, 28, 4982. [Google Scholar] [CrossRef]
- Choi, S.-S.; Lee, S.-H.; Lee, K.-A. A Comparative Study of Hesperetin, Hesperidin and Hesperidin Glucoside: Antioxidant, Anti-Inflammatory, and Antibacterial Activities In Vitro. Antioxidants 2022, 11, 1618. [Google Scholar] [CrossRef]
- Choudhary, H.R.; B., D.K.; Kadiri, S.K.; Khobragade, D.S.; Tiwari, P. Integrating Biosensors in Phytochemical Research: Challenges and Breakthroughs. Recent Patents Biotechnol. 2025, 20, 1–16. [CrossRef]
- Ciriminna, R.; Petri, G.L.; Angellotti, G.; Luque, R.; Tixier, A.F.; Meneguzzo, F.; Pagliaro, M. Citrus Flavonoids as Antimicrobials. Chem. Biodivers. 2025, 22, e202403210. [Google Scholar] [CrossRef]
- Črnivec, I.G.O.; Skrt, M.; Šeremet, D.; Sterniša, M.; Farčnik, D.; Štrumbelj, E.; Poljanšek, A.; Cebin, N.; Pogačnik, L.; Možina, S.S.; et al. Waste streams in onion production: Bioactive compounds, quercetin and use of antimicrobial and antioxidative properties. Waste Manag. 2021, 126, 476–486. [Google Scholar] [CrossRef]
- Dai, X.; Zhang, X.; Chen, W.; Chen, Y.; Zhang, Q.; Mo, S.; Lu, J. Dihydroartemisinin: A Potential Natural Anticancer Drug. Int. J. Biol. Sci. 2021, 17, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Kwon, M.; Kim, J.-Y. Enhancement of specialized metabolites using CRISPR/Cas gene editing technology in medicinal plants. Front. Plant Sci. 2024, 15, 1279738. [Google Scholar] [CrossRef] [PubMed]
- Davidova, S.; Galabov, A.S.; Satchanska, G. Antibacterial, Antifungal, Antiviral Activity, and Mechanisms of Action of Plant Polyphenols. Microorganisms 2024, 12, 2502. [Google Scholar] [CrossRef] [PubMed]
- Cherubim, D.J.d.L.; Martins, C.V.B.; Fariña, L.O.; Lucca, R.A.d.S.d. Polyphenols as natural antioxidants in cosmetics applications. J. Cosmet. Dermatol. 2020, 19, 33–37. [Google Scholar] [CrossRef]
- De, R.; Sarkar, A.; Ghosh, P.; Ganguly, M.; Karmakar, B.C.; Saha, D.R.; Halder, A.; Chowdhury, A.; Mukhopadhyay, A.K. Antimicrobial activity of ellagic acid against Helicobacter pylori isolates from India and during infections in mice. J. Antimicrob. Chemother. 2018, 73, 1595–1603. [Google Scholar] [CrossRef]
- Diab, M.K.; Abu-Elsaoud, A.M.; Salama, M.G.; Ghareeb, E.M. Phytochemical treasure troves—insights into bioactivities, phytochemistry, and uses of Artemisia species. Phytochem. Rev. 2025, 1–38. [Google Scholar] [CrossRef]
- Dibal, N.I.; Garba, S.H.; Jacks, T.W. Acute Toxicity of Quercetin From Onion Skin in Mice. Pharm. Biomed. Res. 2020. [Google Scholar] [CrossRef]
- Ding, Y.; Yu, Y. Therapeutic potential of flavonoids in gastrointestinal cancer: Focus on signaling pathways and improvement strategies (Review). Mol. Med. Rep. 2025, 31, 1–34. [Google Scholar] [CrossRef]
- Du, W.; Zhou, M.; Liu, Z.; Chen, Y.; Li, R. Inhibition effects of low concentrations of epigallocatechin gallate on the biofilm formation and hemolytic activity of Listeria monocytogenes. Food Control. 2018, 85, 119–126. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Scientific opinion on the safety of green tea catechins. EFSA J. 2018, 16, e05239. [CrossRef]
- El-Rayes, T. K., El Basuini, M. F., Elghioushy, A. B., El-Damrawy, S. Z., Mamdouh, M., Taha, A. E., & Abd El-Hack, M. E. Dietary inclusion of Artemisia annua improves antioxidant performance, immunoglobulin protein levels, lipid profile, carcass characteristics, meat quality, and histomorphometric features of broiler chickens. Annals of Animal Science 2025, 25, 225-238. https://www.proquest.com/openview/c14a38224c48e74ccf76f6c2c1b8cb13/1?pq-origsite=gscholar&cbl=1976406.
- Evtyugin, D.D.; Magina, S.; Evtuguin, D.V. Recent Advances in the Production and Applications of Ellagic Acid and Its Derivatives. A Review. Molecules 2020, 25, 2745. [Google Scholar] [CrossRef] [PubMed]
- Faria, W.C.S.; da Silva, A.A.; Veggi, N.; Kawashita, N.H.; Lemes, S.A.d.F.; de Barros, W.M.; Cardoso, E.d.C.; Converti, A.; Moura, W.d.M.; Bragagnolo, N. Acute and subacute oral toxicity assessment of dry encapsulated and non-encapsulated green coffee fruit extracts. J. Food Drug Anal. 2020, 28, 14–161. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.-X.; Hong, J.-X.; Wang, Q.; Fan, Y.-Y.; Yuan, C.-T.; Lei, X.-H.; Zhu, M.; Qin, A.; Chen, H.-X.; Hong, D. Dihydroartemisinin prevents breast cancer-induced osteolysis via inhibiting both breast caner cells and osteoclasts. Sci. Rep. 2016, 6, srep19074. [Google Scholar] [CrossRef]
- de Franco, E.P.D.; Contesini, F.J.; da Silva, B.L.; Fernandes, A.M.A.d.P.; Leme, C.W.; Cirino, J.P.G.; Campos, P.R.B.; Carvalho, P.d.O. Enzyme-assisted modification of flavonoids from Matricaria chamomilla: antioxidant activity and inhibitory effect on digestive enzymes. J. Enzym. Inhib. Med. Chem. 2020, 35, 42–49. [Google Scholar] [CrossRef]
- Fuentes, J.; Arias-Santé, M.F.; Atala, E.; Pastene, E.; Kogan, M.J.; Speisky, H. Low nanomolar concentrations of a quercetin oxidation product, which naturally occurs in onion peel, protect cells against oxidative damage. Food Chem. 2020, 314, 126166. [Google Scholar] [CrossRef]
- Furniturewalla, A.; Barve, K. Approaches to overcome bioavailability inconsistencies of epigallocatechin gallate, a powerful anti-oxidant in green tea. Food Chem. Adv. 2022, 1, 100037. [Google Scholar] [CrossRef]
- Gan, L.; Wang, W.; Jiang, J.; Tian, K.; Liu, W.; Cao, Z. Dual role of Nrf2 signaling in hepatocellular carcinoma: promoting development, immune evasion, and therapeutic challenges. Front. Immunol. 2024, 15, 1429836. [Google Scholar] [CrossRef]
- Gang, G.; Gao, R.; Li, R.; Jin, X.; Xing, Y.; Yan, S.; Xu, Y.; Shi, B. Study on the Regulatory Effect of Water Extract of Artemisia annua L. on Antioxidant Function of Mutton Sheep via the Keap1/Nrf2 Signaling Pathway. Antioxidants 2025, 14, 885. [Google Scholar] [CrossRef]
- Gang, G.; Gao, R.; Zhao, H.; Xu, Y.; Xing, Y.; Jin, X.; Hong, L.; Yan, S.; Shi, B. Effects of water extracts of Artemisia annua L. on rumen immune and antioxidative indexes, fermentation parameters and microbials diversity in lambs. Front. Microbiol. 2024, 15, 1485882. [Google Scholar] [CrossRef]
- Gao, H.; Pei, X.; Song, X.; Wang, S.; Yang, Z.; Zhu, J.; Lin, Q.; Zhu, Q.; Yang, X. Application and development of CRISPR technology in the secondary metabolic pathway of the active ingredients of phytopharmaceuticals. Front. Plant Sci. 2025, 15, 1477894. [Google Scholar] [CrossRef] [PubMed]
- Gavarić, N., Aćimović, M., Kladar, N., Hitl, M., Drljača Lero, J., Milić, N., & Radovanović, K. Unlocking the Bioactivity of Sweet Wormwood (Artemisia annua L., Asteraceae) Ethanolic Extract: Phenolics, Antioxidants, and Cytotoxic Effects. Pharmaceutics 2025, 17, 890. https://www.mdpi.com/1999-4923/17/7/890.
- Grzelak-Błaszczyk, K.; Milala, J.; Kosmala, M.; Kołodziejczyk, K.; Sójka, M.; Czarnecki, A.; Klewicki, R.; Juśkiewicz, J.; Fotschki, B.; Jurgoński, A. Onion quercetin monoglycosides alter microbial activity and increase antioxidant capacity. J. Nutr. Biochem. 2018, 56, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.; Orellana-Palacios, J.C.; Aghababaei, F.; Gonzalez-Serrano, D.J.; Moreno, A.; Lorenzo, J.M. Plant by-product antioxidants: Control of protein-lipid oxidation in meat and meat products. LWT 2022, 169, 114003. [Google Scholar] [CrossRef]
- Halliwell, B. Understanding mechanisms of antioxidant action in health and disease. Nat. Rev. Mol. Cell Biol. 2023, 25, 13–33. [Google Scholar] [CrossRef]
- Hasan, Z.; Islam, A.; Khan, L.A. Spectroscopic investigations on fungal aspartic protease as target of gallic acid. Int. J. Biol. Macromol. 2023, 228, 333–345. [Google Scholar] [CrossRef]
- Hoang, H.T.; Moon, J.-Y.; Lee, Y.-C. Natural Antioxidants from Plant Extracts in Skincare Cosmetics: Recent Applications, Challenges and Perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Huang, Z.; Gan, S.; Zhuang, X.; Chen, Y.; Lu, L.; Wang, Y.; Qi, X.; Feng, Q.; Huang, Q.; Du, B.; et al. Artesunate Inhibits the Cell Growth in Colorectal Cancer by Promoting ROS-Dependent Cell Senescence and Autophagy. Cells 2022, 11, 2472. [Google Scholar] [CrossRef]
- Hussain, Y.; Khan, H.; Alsharif, K.F.; Khan, A.H.; Aschner, M.; Saso, L. The Therapeutic Potential of Kaemferol and Other Naturally Occurring Polyphenols Might Be Modulated by Nrf2-ARE Signaling Pathway: Current Status and Future Direction. Molecules 2022, 27, 4145. [Google Scholar] [CrossRef]
- Ikawa, M.; Okazawa, H.; Yoneda, M. Molecular imaging for mitochondrial metabolism and oxidative stress in mitochondrial diseases and neurodegenerative disorders. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2021, 1865, 129832. [Google Scholar] [CrossRef]
- Jan, R.; Khan, M.; Asaf, S.; Lubna; Asif, S.; Kim, K.-M. Bioactivity and Therapeutic Potential of Kaempferol and Quercetin: New Insights for Plant and Human Health. Plants 2022, 11, 2623. [CrossRef]
- Jia, X.; Wang, L.; Zhao, H.; Zhang, Y.; Chen, Z.; Xu, L.; Yi, K. The origin and evolution of salicylic acid signaling and biosynthesis in plants. Mol. Plant 2023, 16, 245–259. [Google Scholar] [CrossRef]
- Jomova, K., Alomar, S. Y., Valko, R., Liska, J., Nepovimova, E., Kuca, K., & Valko, M. Flavonoids and their role in oxidative stress, inflammation, and human diseases. Chemico-Biological Interactions 2025, 111489. https://www.sciencedirect.com/science/article/pii/S000927972500119X.
- Kandemir, K.; Tomas, M.; McClements, D.J.; Capanoglu, E. Recent advances on the improvement of quercetin bioavailability. Trends Food Sci. Technol. 2022, 119, 192–200. [Google Scholar] [CrossRef]
- Karalija, E.; Macanović, A.; Ibragić, S. Revisiting Traditional Medicinal Plants: Integrating Multiomics, In Vitro Culture, and Elicitation to Unlock Bioactive Potential. Plants 2025, 14, 2029. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.T.B.; Fatima, H.; Naz, I.; Kanwal, N.; Haq, I.-U. Pre-clinical studies comparing the anti-inflammatory potential of artemisinic compounds by targeting NFκB/TNF-α/NLRP3 and Nrf2/TRX pathways in Balb/C mice. Front. Pharmacol. 2024, 15, 1352827. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.; Yehye, W.A.; Etxeberria, A.E.; Alhadi, A.A.; Dezfooli, S.M.; Julkapli, N.B.M.; Basirun, W.J.; Seyfoddin, A. Nanoantioxidants: Recent Trends in Antioxidant Delivery Applications. Antioxidants 2019, 9, 24. [Google Scholar] [CrossRef]
- Khan, A.; Ikram, M.; Hahm, J.R.; Kim, M.O. Antioxidant and Anti-Inflammatory Effects of Citrus Flavonoid Hesperetin: Special Focus on Neurological Disorders. Antioxidants 2020, 9, 609. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Maalik, A.; Murtaza, G. Inhibitory mechanism against oxidative stress of caffeic acid. J. Food Drug Anal. 2016, 24, 695–702. [Google Scholar] [CrossRef]
- Kim, W. S., Choi, W. J., Lee, S., Kim, W. J., Lee, D. C., Sohn, U. D., Shin, H.S., & Kim, W. Anti-inflammatory, antioxidant and antimicrobial effects of artemisinin extracts from Artemisia annua L. The Korean Journal of Physiology and Pharmacology 2015, 19, 21–27.
- Kishi, S.; Nagasu, H.; Kidokoro, K.; Kashihara, N. Oxidative stress and the role of redox signalling in chronic kidney disease. Nat. Rev. Nephrol. 2024, 20, 101–119. [Google Scholar] [CrossRef]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Lipert, A.; Szymańska, G. Yeast Extract Stimulates Ginsenoside Production in Hairy Root Cultures of American Ginseng Cultivated in Shake Flasks and Nutrient Sprinkle Bioreactors. Molecules 2017, 22, 880. [Google Scholar] [CrossRef]
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28, 1359–1390. [Google Scholar] [CrossRef] [PubMed]
- Kuang, M.; Cen, Y.; Qin, R.; Shang, S.; Zhai, Z.; Liu, C.; Pan, X.; Zhou, H. Artesunate Attenuates Pro-Inflammatory Cytokine Release from Macrophages by Inhibiting TLR4-Mediated Autophagic Activation via the TRAF6-Beclin1-PI3KC3 Pathway. Cell. Physiol. Biochem. 2018, 47, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Lala, S. Nanoparticles as elicitors and harvesters of economically important secondary metabolites in higher plants: A review. IET Nanobiotechnology 2021, 15, 28–57. [Google Scholar] [CrossRef] [PubMed]
- Laurindo, L.F.; de Maio, M.C.; Minniti, G.; de Góes Corrêa, N.; Barbalho, S.M.; Quesada, K.; Guiguer, E.L.; Sloan, K.P.; Detregiachi, C.R.P.; Araújo, A.C.; et al. Effects of Medicinal Plants and Phytochemicals in Nrf2 Pathways during Inflammatory Bowel Diseases and Related Colorectal Cancer: A Comprehensive Review. Metabolites 2023, 13, 243. [Google Scholar] [CrossRef]
- Le, B.; Anh, P.T.N.; Kim, J.-E.; Cheng, J.; Yang, S.H. Rice bran fermentation by lactic acid bacteria to enhance antioxidant activities and increase the ferulic acid, ρ-coumaric acid, and γ-oryzanol content. J. Appl. Biol. Chem. 2019, 62, 257–264. [Google Scholar] [CrossRef]
- Lee, R.; Lee, W.-Y.; Kim, D.-W.; Park, H.-J. Artemisinin alleviates cisplatin-induced damage in GC-1 spermatogonia through ER stress mechanisms. Heliyon 2025, 11, e42579. [Google Scholar] [CrossRef]
- Le, N.P.K.; Herz, C.; Gomes, J.V.D.; Förster, N.; Antoniadou, K.; Mittermeier-Kleßinger, V.K.; Mewis, I.; Dawid, C.; Ulrichs, C.; Lamy, E. Comparative Anti-Inflammatory Effects of Salix Cortex Extracts and Acetylsalicylic Acid in SARS-CoV-2 Peptide and LPS-Activated Human In Vitro Systems. Int. J. Mol. Sci. 2021, 22, 6766. [Google Scholar] [CrossRef]
- Lee, E.J.; Patil, B.S.; Yoo, K.S. Antioxidants of 15 onions with white, yellow, and red colors and their relationship with pungency, anthocyanin, and quercetin. LWT-Food Science and Technology 2015, 63, 108–114. [Google Scholar] [CrossRef]
- Les, F.; Prieto, J.M.; Arbonés-Mainar, J.M.; Valero, M.S.; López, V. Bioactive properties of commercialised pomegranate (Punica granatum) juice: antioxidant, antiproliferative and enzyme inhibiting activities. Food Funct. 2015, 6, 2049–2057. [Google Scholar] [CrossRef]
- https://pubs.rsc.org/en/content/articlelanding/2015/fo/c5fo00426h/unauth.
- Li, J.; Yu, Q.; Liu, C.; Zhang, N.; Xu, W. Flavonoids as key players in cold tolerance: molecular insights and applications in horticultural crops. Hortic. Res. 2025, 12, uhae366. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Mabury, S.A. Synthetic Phenolic Antioxidants: A Review of Environmental Occurrence, Fate, Human Exposure, and Toxicity. Environ. Sci. Technol. 2020, 54, 11706–11719. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, S.; Wei, R.; Cui, Z.; Wu, X.; Wei, R.; Xie, L.; Zhou, Y.; Li, W.; Chen, W. Keap1 Cystenine 151 as a Potential Target for Artemisitene-Induced Nrf2 Activation. BioMed Res. Int. 2019, 2019, 5198138. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Lv, Y.; Li, W.; Liao, W.; Jiang, H.; Liu, Y.; Cao, J.; Lu, W.; Feng, Y. Nano-Drug Delivery Systems Based on Natural Products. Int. J. Nanomed. 2024, ume 19, 541–569. [Google Scholar] [CrossRef]
- Maistro, E.L.; Terrazzas, P.M.; Sawaya, A.C.H.F.; Rosa, P.C.P.; Perazzo, F.F.; Gaivão, I.O.d.M. In vivo toxicogenic potential of Salix alba (Salicaceae) bark extract. J. Toxicol. Environ. Heal. Part A 2021, 85, 121–130. [Google Scholar] [CrossRef]
- Mishra, S.; Khushtar, M.; Ahmad, U. Recent Advances in the Pharmacological Potential of Quercetin and Kaempferol: Mechanisms, Therapeutic Applications, and Future Perspectives- A Comprehensive Review. Cuest. de Fisioter. 2025, 54, 5497–5505. [Google Scholar] [CrossRef]
- Mittal, G.; A, P.; Dhali, A.; Prasad, R.; S, Y.; Nurani, K.M.; Găman, M.-A. Plant extracts with antioxidant and hepatoprotective benefits for liver health: A bibliometric analysis of drug delivery systems. World J. Gastroenterol. 2025, 31, 105836. [Google Scholar] [CrossRef]
- Moazzen, A.; Öztinen, N.; Ak-Sakalli, E.; Koşar, M. Structure-antiradical activity relationships of 25 natural antioxidant phenolic compounds from different classes. Heliyon 2022, 8, e10467. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Emwas, A.-H.; Khan, R.A. Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management. Int. J. Mol. Sci. 2023, 24, 5171. [Google Scholar] [CrossRef]
- Mohammed, M. J., Anand, U., Altemimi, A. B., Tripathi, V., Guo, Y., & Pratap-Singh, A. Phenolic composition, antioxidant capacity and antibacterial activity of white wormwood (Artemisia herba-alba). Plants 2021, 10, 164.
- Molaei, E.; Molaei, A.; Abedi, F.; Hayes, A.W.; Karimi, G. Nephroprotective activity of natural products against chemical toxicants: The role of Nrf2/ARE signaling pathway. Food Sci. Nutr. 2021, 9, 3362–3384. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.M.; Wang, J.; Lin, Q.; Ferreira, P.; Avery, M.A.; Elokely, K.; Staines, H.M.; Krishna, S. Selective Inhibition of Plasmodium falciparum ATPase 6 by Artemisinins and Identification of New Classes of Inhibitors after Expression in Yeast. Antimicrob. Agents Chemother. 2022, 66, e0207921. [Google Scholar] [CrossRef] [PubMed]
- Morua, E. , Cuyas, L., & Matías-Hernández, L. The Beneficial Use of Artemisia annua, Artemisinin, and Other Compounds in Animal Health. Animals 2025, 15, 1359. [Google Scholar]
- Mude, H.; Maroju, P.A.; Balapure, A.; Ganesan, R.; Dutta, J.R. Water-soluble caffeic acid-dopamine acid-base complex exhibits enhanced bactericidal, antioxidant, and anticancer properties. Food Chem. 2022, 374, 131830. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: an overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Nair, B.; Menon, A.; Kalidas, M.R.; Nath, L.R.; Calina, D.; Sharifi-Rad, J. Modulating the JAK/STAT pathway with natural products: potential and challenges in cancer therapy. Discov. Oncol. 2025, 16, 1–23. [Google Scholar] [CrossRef]
- Nam, D.-G.; Kim, M.; Choi, A.-J.; Choe, J.-S. Health Benefits of Antioxidant Bioactive Compounds in Ginger (Zingiber officinale) Leaves by Network Pharmacology Analysis Combined with Experimental Validation. Antioxidants 2024, 13, 652. [Google Scholar] [CrossRef]
- Naraki, K.; Rameshrad, M.; Hosseinzadeh, H. Protective effects and therapeutic applications of ellagic acid against natural and synthetic toxicants: A review article. Iranian journal of basic medical sciences 2022, 25, 1402. [Google Scholar] [CrossRef]
- Nuryana, I., Ratnakomala, S., Fahrurrozi, A. B. J., Andriani, A., Putra, F. J. N., Rezamela, E., Wulansari, R., Prawira-Atmaja, M.I., & Lisdiyanti, P. Catechin contents, antioxidant and antibacterial activities of different types of Indonesian tea (Camellia sinensis). Ann. Bogor 2020, 24, 107. https://www.researchgate.net/profile/Isa-Nuryana/publication/348264057_Catechin_Contents_Antioxidant_and_Antibacterial_Activities_of_Different_Types_of_Indonesian_Tea_Camellia_sinensis/links/5ff55a90299bf1408874f73c/Catechin-Contents-Antioxidant-and-Antibacterial-Activities-of-Different-Types-of-Indonesian-Tea-Camellia-sinensis.pdf.
- Nwozo, O.S.; Effiong, E.M.; Aja, P.M.; Awuchi, C.G. Antioxidant, phytochemical, and therapeutic properties of medicinal plants: a review. Int. J. Food Prop. 2023, 26, 359–388. [Google Scholar] [CrossRef]
- Osonwa, U.E.; Hu, M. Bioavailability and Pharmacokinetics of Dihydroartemisinin (DHA) and its Analogs—Mechanistic Studies on its ADME. Curr. Pharmacol. Rep. 2018, 4, 33–44. [Google Scholar] [CrossRef]
- Pan, L.; Wang, Q.; Li, W.; Dong, X.; Xie, S.; Tan, B.; Liu, H.; Wang, Y. Artemisinin supplementation in concentrated cottonseed protein basal diets enhances growth, antioxidant capacity, intestinal immunity and microbiota in hybrid grouper (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂). Aquac. Rep. 2025, 41, 102659. [Google Scholar] [CrossRef]
- Pavlova, E.L.; Zografov, N.N.; Simeonova, L.S. Comparative study on the antioxidant capacities of synthetic influenza inhibitors and ellagic acid in model systems. Biomed. Pharmacother. 2016, 83, 755–762. [Google Scholar] [CrossRef]
- https://www.sciencedirect.com/science/article/abs/pii/S0753332216306886.
- Pawłowska, M.; Nuszkiewicz, J.; Jarek, D.J.; Woźniak, A. Ferroptosis and Metabolic Dysregulation: Emerging Chemical Targets in Cancer and Infection. Molecules 2025, 30, 3020. [Google Scholar] [CrossRef]
- Perez-Ternero, C.; Werner, C.M.; Nickel, A.G.; Herrera, M.D.; Motilva, M.-J.; Böhm, M.; de Sotomayor, M.A.; Laufs, U. Ferulic acid, a bioactive component of rice bran, improves oxidative stress and mitochondrial biogenesis and dynamics in mice and in human mononuclear cells. J. Nutr. Biochem. 2017, 48, 51–61. [Google Scholar] [CrossRef]
- Parveen, B.; Rajinikanth, V.; Narayanan, M. Natural plant antioxidants for food preservation and emerging trends in nutraceutical applications. Discov. Appl. Sci. 2025, 7, 845. [Google Scholar] [CrossRef]
- Patel, J.; Roy, H.; Chintamaneni, P.K.; Patel, R.; Bohara, R. Advanced Strategies in Enhancing the Hepatoprotective Efficacy of Natural Products: Integrating Nanotechnology, Genomics, and Mechanistic Insights. ACS Biomater. Sci. Eng. 2025, 11, 2528–2549. [Google Scholar] [CrossRef]
- Pham, D.-C.; Shibu, M.A.; Mahalakshmi, B.; Velmurugan, B.K. Effects of phytochemicals on cellular signaling: reviewing their recent usage approaches. Crit. Rev. Food Sci. Nutr. 2020, 60, 3522–3546. [Google Scholar] [CrossRef]
- Piątczak, E., Dybowska, M., Płuciennik, E., Kośla, K., Kolniak-Ostek, J., & Kalinowska-Lis, U. Identification and accumulation of phenolic compounds in the leaves and bark of Salix alba (L.) and their biological potential. Biomolecules 2020, 10, 1391.
- Pillai, R.; Hayashi, M.; Zavitsanou, A.-M.; Papagiannakopoulos, T. NRF2: KEAPing Tumors Protected. Cancer Discov. 2022, 12, 625–643. [Google Scholar] [CrossRef]
- Pooja, G.; Shweta, S.; Patel, P. Oxidative stress and free radicals in disease pathogenesis: a review. Discov. Med. 2025, 2, 104. [Google Scholar] [CrossRef]
- Quecan, B.X.V.; Santos, J.T.C.; Rivera, M.L.C.; Hassimotto, N.M.A.; Almeida, F.A.; Pinto, U.M. Effect of Quercetin Rich Onion Extracts on Bacterial Quorum Sensing. Front. Microbiol. 2019, 10, 867. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Rahaman, S.; Islam, R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef] [PubMed]
- Randjelović, P.; Veljković, S.; Stojiljković, N.; Sokolović, D.; Ilić, I.; Laketić, D.; Randjelović, D.; Randjelović, N. The Beneficial Biological Properties of Salicylic Acid. Acta Fac. Medicae Naissensis 2015, 32, 259–265. [Google Scholar] [CrossRef]
- Rasheed, F.; Naeem, M.; Uddin, M.; Khan, M.M.A. Synergistic Effect of Irradiated Sodium Alginate and Methyl Jasmonate on Anticancer Alkaloids Production in Periwinkle (Catharanthus roseus L.). J. Funct. Environ. Bot. 2017, 7, 77–91. [Google Scholar] [CrossRef]
- Rasheed, A.; Kamran, S.H.; Hameed, M.; Siddique, F.; Latif, S.; Bibi, M.; Rizwan, R. Syringic acid loaded chitosan nanoparticles mitigate glycation associated oxidative stress and inflammation in hyperglycaemic rat model. Sci. Rep. 2025, 15, 22778. [Google Scholar] [CrossRef]
- Rithichai, P.; Jirakiattikul, Y.; Nambuddee, R.; Itharat, A.; Addi, M. Effect of Salicylic Acid Foliar Application on Bioactive Compounds and Antioxidant Activity in Holy Basil (Ocimum sanctum L.). Int. J. Agron. 2024, 2024, 8159886. [Google Scholar] [CrossRef]
- Rudrapal, M.; Rakshit, G.; Singh, R.P.; Garse, S.; Khan, J.; Chakraborty, S. Dietary Polyphenols: Review on Chemistry/Sources, Bioavailability/Metabolism, Antioxidant Effects, and Their Role in Disease Management. Antioxidants 2024, 13, 429. [Google Scholar] [CrossRef]
- Sah, A.; Naseef, P.P.; Kuruniyan, M.S.; Jain, G.K.; Zakir, F.; Aggarwal, G. A Comprehensive Study of Therapeutic Applications of Chamomile. Pharmaceuticals 2022, 15, 1284. [Google Scholar] [CrossRef]
- Saikia, L.; Talukdar, N.C.; Dutta, P.P. Exploring the Therapeutic Role of Flavonoids Through AMPK Activation in Metabolic Syndrome: A Narrative Review. Phytotherapy Res. 2025, 39, 1403–1421. [Google Scholar] [CrossRef]
- Salehi, B., Venditti, A., Sharifi-Rad, M., Kręgiel, D., Sharifi-Rad, J., Durazzo, A., Lucarini, M., Santini, A., Souto, E.B., Novellino, E. and Antolak, H. & Martins, N. The therapeutic potential of apigenin. International journal of molecular sciences 2019, 20, 1305.
- Salehi, H.; Rad, A.C.; Lucini, L. Different Forms of Manganese Provide Distinctive Metabolomics Signatures and Bioactive Profiles in Artemisia annua. Phytochem. Anal. 2025. [CrossRef]
- Seema, B.R.; Jyothi, Y.; Bhovi, C.N.; Aradhya, M.V.; Lekhak, M.; Mane, S.R. Advanced up and down methodology for acute toxicity assessment with reliable LD50 verified by aqueous extract of curly kale using wistar rats. Res. J. Pharm. Technol. 2023, 16, 4519–4524. [Google Scholar] [CrossRef]
- A Shalaby, E.; Aboul-Enein, A.; El-Shemy, H.; Abou-Elella, F.; Nassrallah, A. Active Ingredients of Some Egyptian Plants Species with Anticancer Activity and Its Mode of Action. Arab. J. Biotechnol. 2025, 24, 18–40. [Google Scholar] [CrossRef]
- Sharma, N.; Biswas, S.; Al-Dayan, N.; Alhegaili, A.S.; Sarwat, M. Antioxidant Role of Kaempferol in Prevention of Hepatocellular Carcinoma. Antioxidants 2021, 10, 1419. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, W.; Yang, Z.; Subbiah, V.; Suleria, H.A.R. Extraction and characterization of phenolic compounds and their potential antioxidant activities. Environ. Sci. Pollut. Res. 2022, 29, 81112–81129. [Google Scholar] [CrossRef]
- Shi, M.; Zhang, S.; Zheng, Z.; Maoz, I.; Zhang, L.; Kai, G. Molecular regulation of the key specialized metabolism pathways in medicinal plants. J. Integr. Plant Biol. 2024, 66, 510–531. [Google Scholar] [CrossRef]
- Shin, J.M.; Son, Y.-J.; Ha, I.J.; Erdenebileg, S.; Jung, D.S.; Song, D.-G.; Kim, Y.S.; Kim, S.M.; Nho, C.W. Artemisia argyi extract alleviates inflammation in a DSS-induced colitis mouse model and enhances immunomodulatory effects in lymphoid tissues. BMC Complement. Med. Ther. 2022, 22, 64. [Google Scholar] [CrossRef]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiaq, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene lactone! a promising antioxidant, anticancer and moderate antinociceptive agent from Artemisia macrocephala jacquem. BMC Complement. Altern. Med. 2017, 17, 27. [Google Scholar] [CrossRef]
- Shrivastava, S., Uthra, C., Reshi, M., Singh, A., Yadav, D., & Shukla, S. Protective effect of hesperetin against acrylamide induced acute toxicity in rats. Indian J Exp Biol 2018, 56, 164-170. https://d1wqtxts1xzle7.cloudfront.net/77012929/IJEB_20563_20164-170-libre.pdf?1640147462=&response-content-disposition=inline%3B+filename%3DProtective_effect_of_hesperetin_against.pdf&Expires=1754574984&Signature=APPax-hgWB6r1AiJzTJ5bccvN08HKwRkIbD3j50LouNyo6YtD50syth70PPCJvzuqjeVOtbXuUrXWL~HnJhHnhp5xJZCE8Le3NRZdyi7i~guoCIR~611SltGk3yRXmZkL3AE-hutMM6yC9AxehtF64mh37WXn4PWKsgVpVgHJzdxdJfqqwyvsOuY~f5GbUh6a0pz3i6yPRaA8BWmBrOfHwAOtgEZU8mpuKh3Dilf2YgkZao5nWZzLHUD1D2r95ima1ghZKXqK3JgaVtThqgPMgpk6p3aGOaUGIBhtI0Xia3by~fKsypTHrrn1QckoB1oYTIImais82eH1pwFfbHCHg__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA.
- Siddiqui, M. F. M. F., Waghmare, S. P., Hajare, S. W., Deshmukh, R. I. S., & Ali, S. C. S. A. Phytochemical analysis and acute toxicity studies of Artemisia annua in Swiss albino mice. Journal of Pharmacognosy and Phytochemistry 2018, 7, 1893-1895. https://www.researchgate.net/profile/Sharad-Chepte/publication/327882389_Phytochemical_analysis_and_acute_toxicity_studies_of_Artemisia_annua_in_Swiss_albino_mice/links/5bab119092851ca9ed25ec4c/Phytochemical-analysis-and-acute-toxicity-studies-of-Artemisia-annua-in-Swiss-albino-mice.pdf.
- Siddiquee, R.; Mahmood, T.; Ansari, V.A.; Ahsan, F.; Bano, S.; Ahmad, S. Apigenin unveiled: an encyclopedic review of its preclinical and clinical insights. Discov. Plants 2025, 2, 11. [Google Scholar] [CrossRef]
- Singh, J., & Kaur, S. Phyllanthus embilica leaves extract: a potential amylase enzyme inhibitor with antioxidant and antimicrobial activity. Int J Pharma Res 2015, 9, 200–206.
- Silva, J.D.R.; Arruda, H.S.; Andrade, A.C.; Berilli, P.; Borsoi, F.T.; Monroy, Y.M.; Rodrigues, M.V.N.; Sampaio, K.A.; Pastore, G.M.; Junior, M.R.M. Eugenia calycina and Eugenia stigmatosa as Promising Sources of Antioxidant Phenolic Compounds. Plants 2024, 13, 2039. [Google Scholar] [CrossRef]
- Singh, A.; Singh, J.; Parween, G.; Khator, R.; Monga, V. A comprehensive review of apigenin a dietary flavonoid: biological sources, nutraceutical prospects, chemistry and pharmacological insights and health benefits. Crit. Rev. Food Sci. Nutr. 2024, 1–37. [Google Scholar] [CrossRef]
- Song, C.-H.; Kim, N.; Nam, R.H.; Choi, S.I.; Kang, C.; Jang, J.Y.; Nho, H.; Shin, E.; Lee, H.-N. Nuclear Factor Erythroid 2-related Factor 2 Knockout Suppresses the Development of Aggressive Colorectal Cancer Formation Induced by Azoxymethane/Dextran Sulfate Sodium-Treatment in Female Mice. J. Cancer Prev. 2021, 26, 41. [Google Scholar] [CrossRef]
- Jeong, D.E.; Song, H.J.; Lim, S.; Lee, S.J.; Lim, J.E.; Nam, D.-H.; Joo, K.M.; Jeong, B.C.; Jeon, S.S.; Choi, H.Y.; et al. Repurposing the anti-malarial drug artesunate as a novel therapeutic agent for metastatic renal cell carcinoma due to its attenuation of tumor growth, metastasis, and angiogenesis. Oncotarget 2015, 6, 33046. [Google Scholar] [CrossRef]
- Sousa, C., Gabriel, C., Cerqueira, F., Manso, M. C., & Vinha, A. F. (2015). Coffee industrial waste as a natural source of bioactive compounds with antibacterial and antifungal activities. https://core.ac.uk/reader/301338150.
- Srinivasulu, C., Ramgopal, M., Ramanjaneyulu, G., Anuradha, C. M., & Kumar, C. S. Syringic acid (SA)‒a review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomedicine & Pharmacotherapy 2018, 108, 547–557.
- Sultana, R.; Imam, Z.; Kumar, R.R.; Banu, V.S.; Nahakpam, S.; Bharti, R.; Bharadwaj, C.; Singh, A.K.; Pasala, R.K.; Singh, D.R.; et al. Signaling and Defence Mechanism of Jasmonic and Salicylic Acid Response in Pulse Crops: Role of WRKY Transcription Factors in Stress Response. J. Plant Growth Regul. 2025, 44, 5–21. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Sun, J.; Xufeng, J.; Qiu, Z.; Gao, Q.; Zhu, Z.; He, J.; Gao, S.; Sui, X. Artemisiae Annuae Herba: from anti-malarial legacy to emerging anti-cancer potential. Theranostics 2025, 15, 7346. [Google Scholar] [CrossRef] [PubMed]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali; Kepel, B.J.; Idroes, R.; Effendi, Y.; Alam Sakib, S.; Bin Emran, T.; Riganti, C. Potential of Plant Bioactive Compounds as SARS-CoV-2 Main Protease (Mpro) and Spike (S) Glycoprotein Inhibitors: A Molecular Docking Study. Scientifica 2020, 2020, 6307457. [CrossRef]
- Thanikachalam, P.V.; Devaraji, M.; Jayaprakash, N.; Sivakumar, H.; Balamurugan, D.; Shanmugan, H.K. Phytochemical innovations in oral cancer therapy: targeting oncogenic pathways with natural compounds. J. Complement. Integr. Med. 2025. [Google Scholar] [CrossRef] [PubMed]
- Pojskic, L.; Tomic, N.; Kalajdzic, A.; Ramic, J.; Kadric, N.L.; Ikanovic, T.; Maksimovic, M.; Pojskic, N. Screening of preferential binding affinity of selected natural compounds to SARS-CoV-2 proteins using in silico methods. Eurasian J. Med. Oncol. 2020, 4, 319–323. [Google Scholar] [CrossRef]
- Trifan, A.; Zengin, G.; Sinan, K.I.; Sieniawska, E.; Sawicki, R.; Maciejewska-Turska, M.; Skalikca-Woźniak, K.; Luca, S.V. Unveiling the Phytochemical Profile and Biological Potential of Five Artemisia Species. Antioxidants 2022, 11, 1017. [Google Scholar] [CrossRef]
- Vaiserman, A.; Koliada, A.; Zayachkivska, A.; Lushchak, O. Nanodelivery of Natural Antioxidants: An Anti-aging Perspective. Front. Bioeng. Biotechnol. 2020, 7, 447. [Google Scholar] [CrossRef]
- Velumani, K.; Rajan, P.S.; Shaik, M.R.; Hussain, S.A.; Shaik, B.; Guru, A.; Issac, P.K. Protective Effect of Artemisinin Against Luperox Induced Oxidative Stress and Insulin Resistance via Pi3k/Akt Pathway in Zebrafish Larvae. Cell Biochem. Biophys. 2025, 1–13. [Google Scholar] [CrossRef]
- Verotta, L.; Panzella, L.; Antenucci, S.; Calvenzani, V.; Tomay, F.; Petroni, K.; Caneva, E.; Napolitano, A. Fermented pomegranate wastes as sustainable source of ellagic acid: Antioxidant properties, anti-inflammatory action, and controlled release under simulated digestion conditions. Food Chem. 2018, 246, 129–136. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.; Wan, X.; Yang, C.S.; Zhang, J. Green tea polyphenol (−)-epigallocatechin-3-gallate triggered hepatotoxicity in mice: Responses of major antioxidant enzymes and the Nrf2 rescue pathway. Toxicol. Appl. Pharmacol. 2015, 283, 65–74. [Google Scholar] [CrossRef]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, S.; Li, R.; Zhao, Q. Unraveling the specialized metabolic pathways in medicinal plant genomes: a review. Front. Plant Sci. 2024, 15, 1459533. [Google Scholar] [CrossRef]
- Wdowiak, K.; Walkowiak, J.; Pietrzak, R.; Bazan-Woźniak, A.; Cielecka-Piontek, J. Bioavailability of Hesperidin and Its Aglycone Hesperetin—Compounds Found in Citrus Fruits as a Parameter Conditioning the Pro-Health Potential (Neuroprotective and Antidiabetic Activity)—Mini-Review. Nutrients 2022, 14, 2647. [Google Scholar] [CrossRef]
- Wei, T.; Liu, J. Anti-angiogenic properties of artemisinin derivatives (Review). Int. J. Mol. Med. 2017, 40, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Li, Z.; Zhang, Y.; Wu, H.; Zhang, T.; Wang, W. Artemisinin and its derivatives as promising therapies for autoimmune diseases. Heliyon 2024, 10, e27972. [Google Scholar] [CrossRef] [PubMed]
- Xin, T. A. O., & Tian, W. A. N. G. In vitro and in vivo antioxidant activities of three major polyphenolic compounds in pomegranate peel: Ellagic acid, punicalin, and punicalagin. Journal of integrative agriculture 2017, 16, 1808–1818.
- Yang, Y.; Liu, M.; Liu, C.; Tang, S.; Gu, D.; Tian, J.; Huang, D.; He, F. Ellagic acid from pomegranate peel: Consecutive countercurrent chromatographic separation and antioxidant effect. Biomed. Chromatogr. 2023, 37, e5662. [Google Scholar] [CrossRef]
- Zduńska, K., Dana, A., Kolodziejczak, A., & Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin pharmacology and physiology 2018, 31, 332–336.
- Zhao, S.; Nan, Y.; Yao, R.; Wang, L.; Zeng, X.; Aadil, R.M.; Shabbir, M.A. Antibacterial Activity and Transcriptomic Analysis of Hesperetin against Alicyclobacillus acidoterrestris Vegetative Cells. Foods 2023, 12, 3276. [Google Scholar] [CrossRef]
- Zhao, Z.; Yang, Q.; Sun, Y.; Ruan, X. Unveiling the antioxidant and anti-inflammatory potential of syringic acid: mechanistic insights and pathway interactions. Front. Pharmacol. 2025, 16, 1615294. [Google Scholar] [CrossRef]
- Zheng, L.; Jacquier, J.-C.; Harbourne, N. Preparation of Polyphenol-Rich Herbal Beverages from White Willow (Salix alba) Bark with Potential Alzheimer’s Disease Inhibitory Activity In Silico. Beverages 2024, 10, 75. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, Y.; Wang, S.; Wang, K.; Tao, L.; Zou, M.; Chen, N.; Xu, J.; Liu, S.; Li, X. Artesunate suppresses RANKL-induced osteoclastogenesis through inhibition of PLCγ1-Ca 2+ –NFATc1 signaling pathway and prevents ovariectomy-induced bone loss. Biochem. Pharmacol. 2017, 124, 57–68. [Google Scholar] [CrossRef]
- Zhuang, W.-B.; Li, Y.-H.; Shu, X.-C.; Pu, Y.-T.; Wang, X.-J.; Wang, T.; Wang, Z. The Classification, Molecular Structure and Biological Biosynthesis of Flavonoids, and Their Roles in Biotic and Abiotic Stresses. Molecules 2023, 28, 3599. [Google Scholar] [CrossRef]
- Zhou, F.; Li, G.; Tan, R.; Wu, G.; Deng, C. Artemisinins in autoimmune diseases: effects and mechanisms in systemic lupus erythematosus and rheumatoid arthritis. Br. J. Pharmacol. 2025, 182, 3411–3427. [Google Scholar] [CrossRef]
- Zufarov, O., & Serkayev, K. Natural antioxidants. Technical science and innovation 2024, 2024, 19–25.
Figure 1.
Natural Antioxidant Arsenal: Key Bioactive Compounds from Diverse Sources.

Figure 2.
Schematic Artemisinin-Induced Nrf2/Keap1 Signaling Pathway Activation Enhances Antioxidant Gene Expression in Cells (Modified from Gang et al., 2025).
Figure 2.
Schematic Artemisinin-Induced Nrf2/Keap1 Signaling Pathway Activation Enhances Antioxidant Gene Expression in Cells (Modified from Gang et al., 2025).
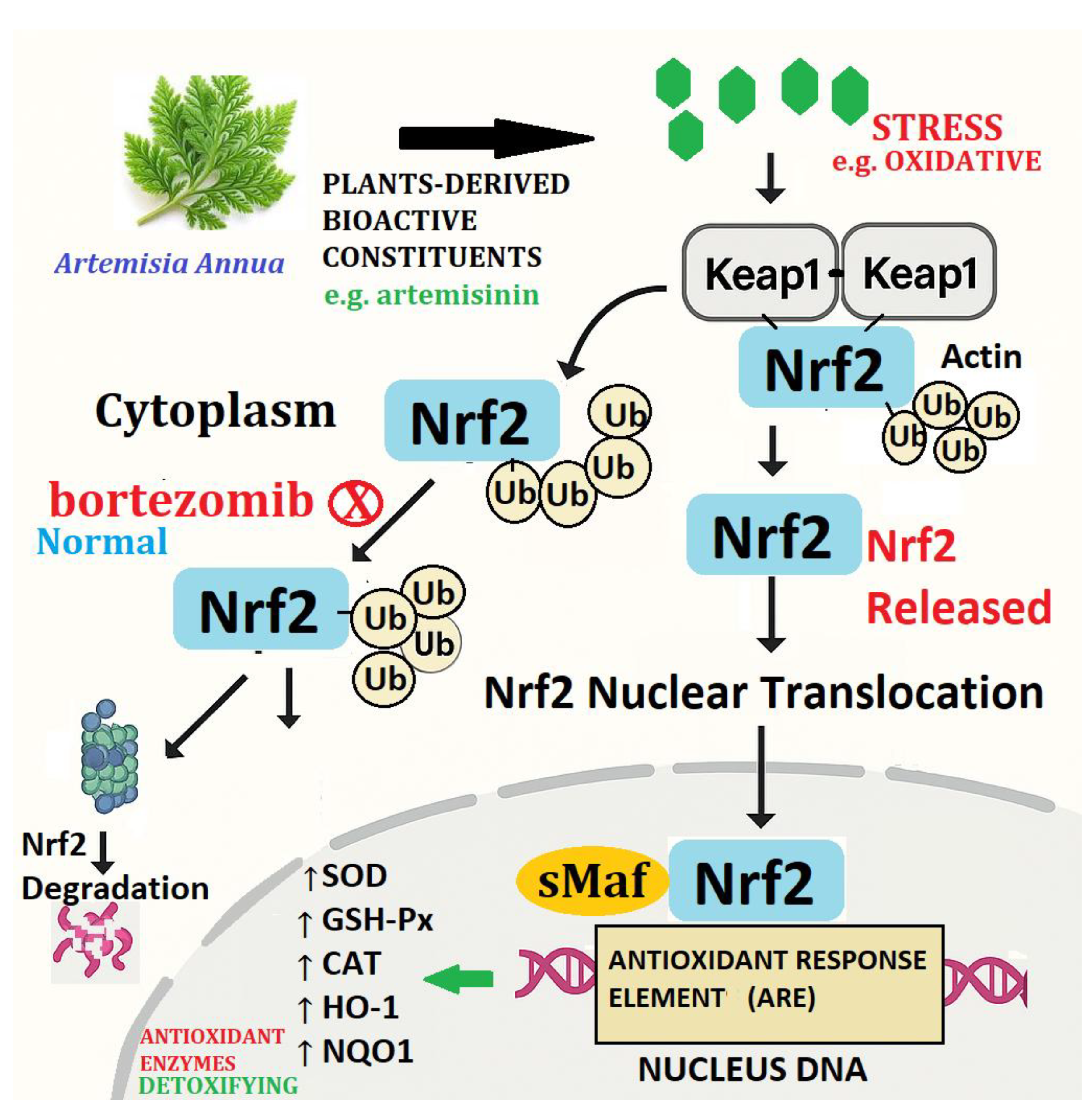
Figure 3.
Bioactive Pathways of Natural Antioxidants. Note: CYP450 = Cytochrome P450, MAPKs = Mitogen-Activated Protein Kinases, Nrf2 = Nuclear factor erythroid 2–related factor 2, NF-κB = Nuclear Factor kappa-light-chain-enhancer of activated B cells, COX-2 = Cyclooxygenase-2, TNF-α = Tumor Necrosis Factor-alpha, Wnt/β-catenin = Wnt signaling pathway involving β-catenin, Keap1 = Kelch-like ECH-associated protein 1, LasR = Quorum sensing transcriptional regulator in Pseudomonas aeruginosa, Rin-5F = Rat insulinoma cell line, HepG2 = Human hepatocellular carcinoma cell line. Line colour: Blue = phenols, Chocolate = flavonoids, Orange = artemisinin, Blue+Orange = combined phenols and possible artemisinin action.
Figure 3.
Bioactive Pathways of Natural Antioxidants. Note: CYP450 = Cytochrome P450, MAPKs = Mitogen-Activated Protein Kinases, Nrf2 = Nuclear factor erythroid 2–related factor 2, NF-κB = Nuclear Factor kappa-light-chain-enhancer of activated B cells, COX-2 = Cyclooxygenase-2, TNF-α = Tumor Necrosis Factor-alpha, Wnt/β-catenin = Wnt signaling pathway involving β-catenin, Keap1 = Kelch-like ECH-associated protein 1, LasR = Quorum sensing transcriptional regulator in Pseudomonas aeruginosa, Rin-5F = Rat insulinoma cell line, HepG2 = Human hepatocellular carcinoma cell line. Line colour: Blue = phenols, Chocolate = flavonoids, Orange = artemisinin, Blue+Orange = combined phenols and possible artemisinin action.
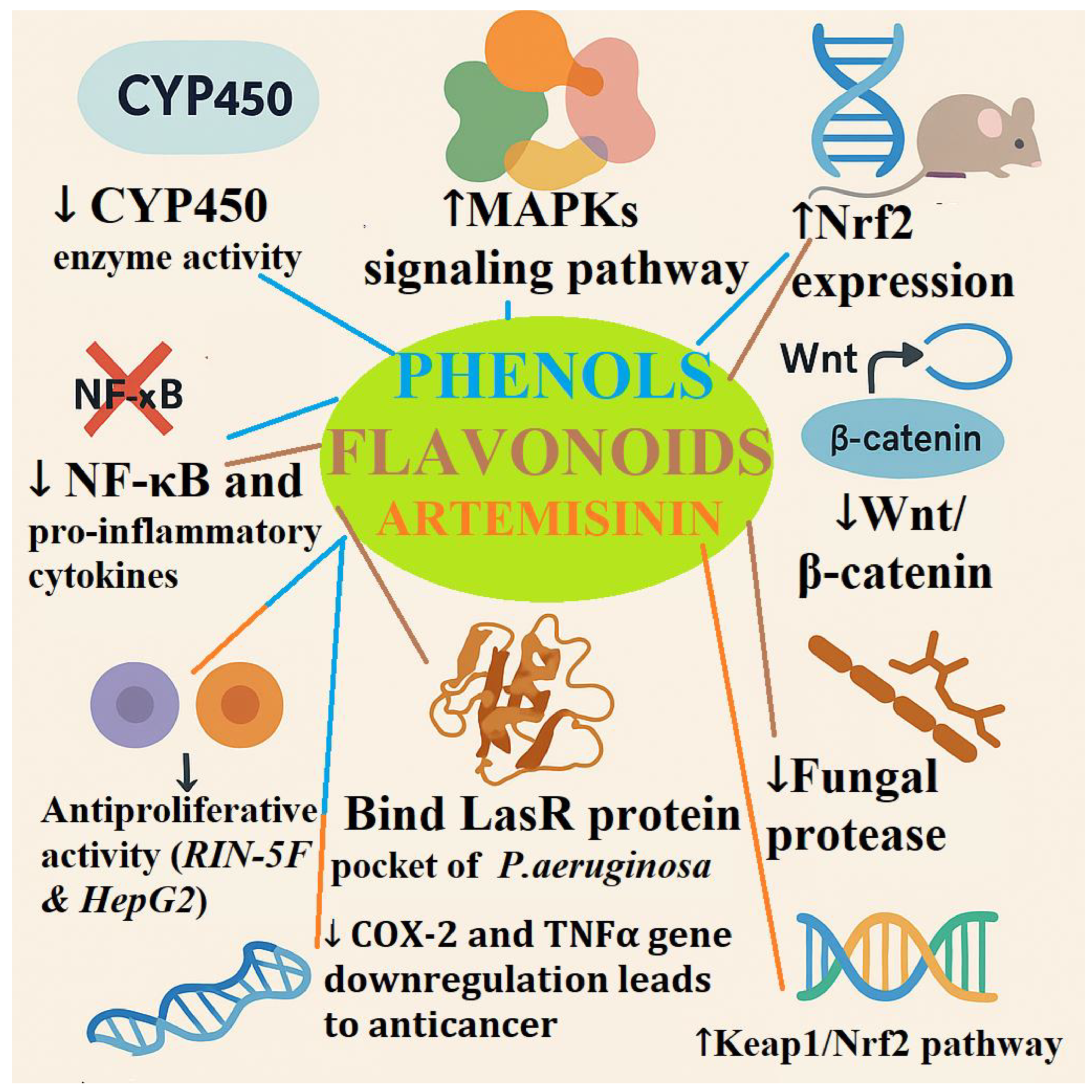
Figure 4.
PRISMA 2020 Flow Diagram for Study Selection in Antioxidant Research (2015–2025)(Haddaway, N. R., Page, M. J., Pritchard, C. C., & McGuinness, L. A. (2022). PRISMA2020: An R package and Shiny app for producing PRISMA 2020-compliant flow diagrams, with interactivity for optimised digital transparency and Open Synthesis Campbell Systematic Reviews, 18, e1230. https://doi.org/10.1002/cl2.1230).
Figure 4.
PRISMA 2020 Flow Diagram for Study Selection in Antioxidant Research (2015–2025)(Haddaway, N. R., Page, M. J., Pritchard, C. C., & McGuinness, L. A. (2022). PRISMA2020: An R package and Shiny app for producing PRISMA 2020-compliant flow diagrams, with interactivity for optimised digital transparency and Open Synthesis Campbell Systematic Reviews, 18, e1230. https://doi.org/10.1002/cl2.1230).
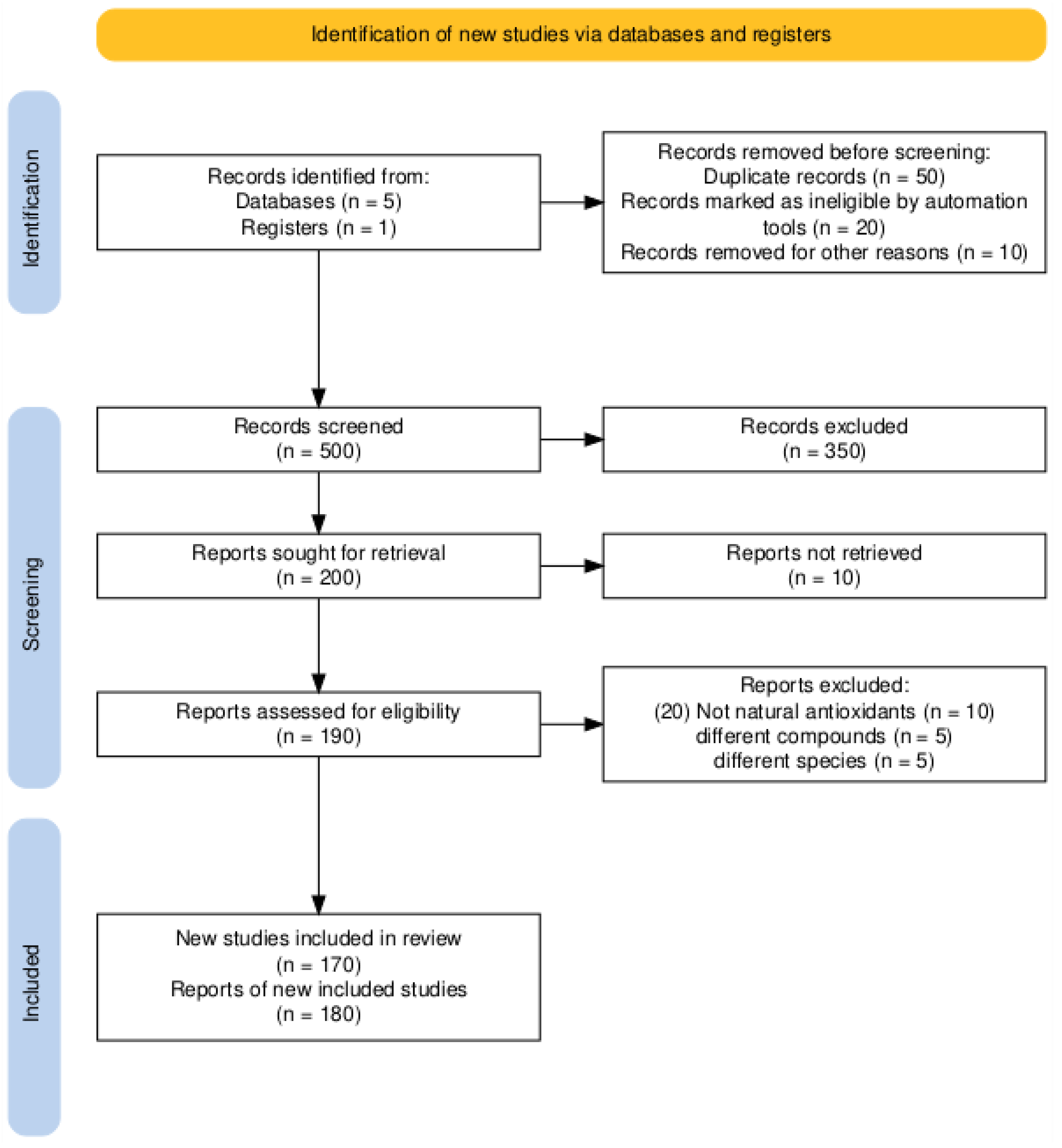
Figure 5.
Selected Phenolic Acids in Gene Regulation: Anti-inflammatory, Antioxidant, and Cytoprotective Outcomes from Molecular Target Studies.
Figure 5.
Selected Phenolic Acids in Gene Regulation: Anti-inflammatory, Antioxidant, and Cytoprotective Outcomes from Molecular Target Studies.
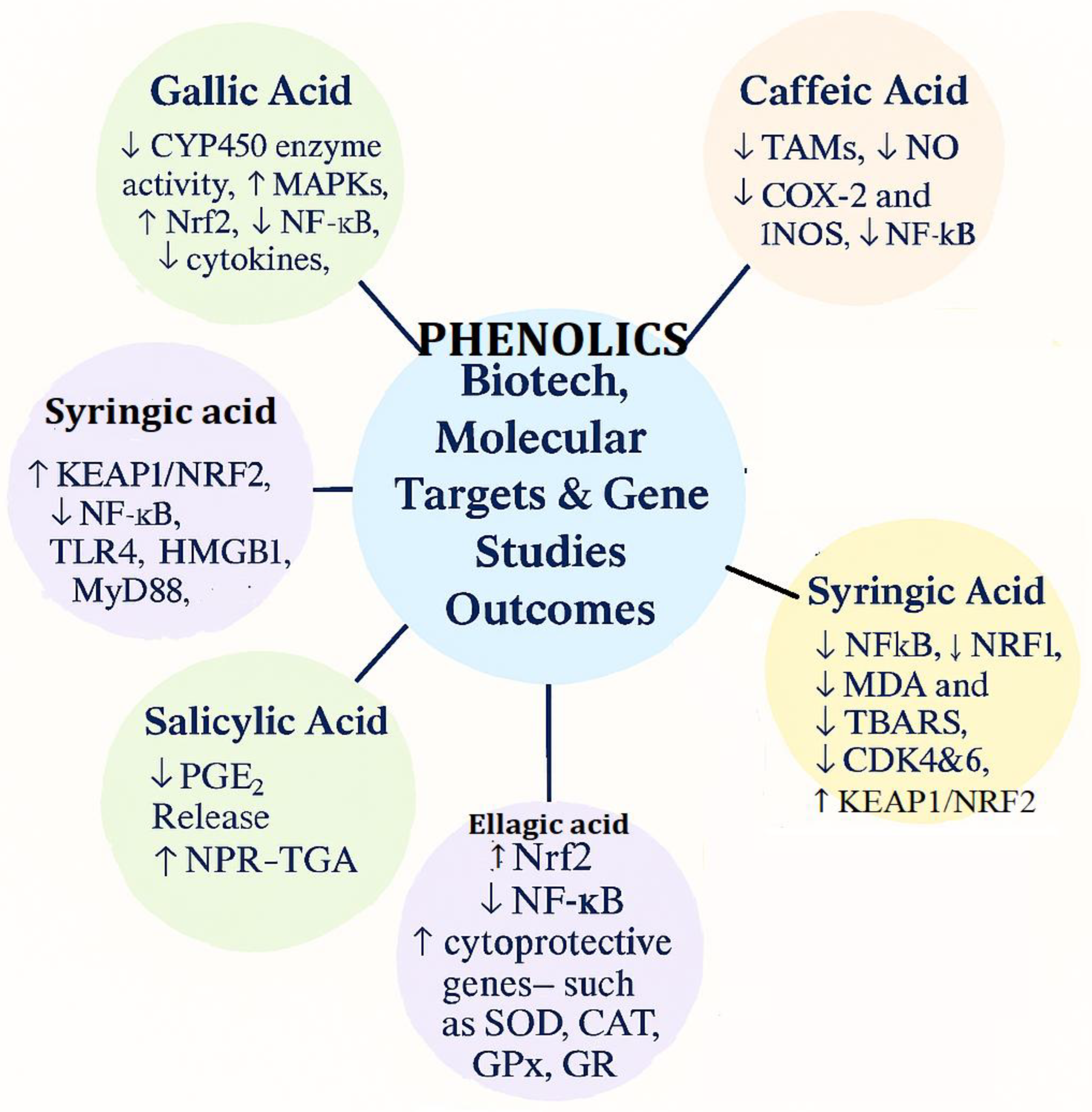
Figure 6.
Selected Flavonoids in Biotechnological and Gene Pathway Modulation; Gene Regulatory Roles in Antioxidant, Neuroprotective, Antiviral, and Metabolic Pathways. Note: LasR = Pseudomonas aeruginosa quorum-sensing regulator, NF-κB = Nuclear factor kappa-light-chain-enhancer of activated B cells, PI3K = Phosphoinositide 3-kinase, Akt = Protein kinase B, Nrf2 = Nuclear factor erythroid 2–related factor 2, MDA = Malondialdehyde, GFAP = Glial fibrillary acidic protein, Iba-1 = Ionized calcium-binding adapter molecule 1, ERK1/2 = Extracellular signal-regulated kinases 1/2, TNF-α = Tumor necrosis factor-alpha, GABA = Gamma-aminobutyric acid, BDNF = Brain-derived neurotrophic factor, JNK = c-Jun N-terminal kinase, MAPKs = Mitogen-activated protein kinases, PLpro = Papain-like protease, 3CLpro = 3-chymotrypsin-like protease, Main protease = SARS-CoV-2 main viral protease, ACE2 = Angiotensin-converting enzyme 2, TJP = Tight junction proteins, JAM-2 = Junctional adhesion molecule 2, MUC-2 = Mucin 2, HO-1 = Heme oxygenase-1, SOD = Superoxide dismutase, GSH = Reduced glutathione, GSSG = Oxidized glutathione, p53 = Tumor suppressor protein, PTEN = Phosphatase and tensin homolog, p21 = Cyclin-dependent kinase inhibitor, STAT3 = Signal transducer and activator of transcription 3, EGFR = Epidermal growth factor receptor, Wnt = Wingless-related integration site, P-gp = P-glycoprotein. Arrow line colour: red line=Quercetine, blue line=Kaempferol, green line=Apigenin, chocolateline line=Hesperetin, Yellow line=EGCG(Epigallocatechin Gallate), ↓= Inhibits or downregulates, ↑= upregulates.
Figure 6.
Selected Flavonoids in Biotechnological and Gene Pathway Modulation; Gene Regulatory Roles in Antioxidant, Neuroprotective, Antiviral, and Metabolic Pathways. Note: LasR = Pseudomonas aeruginosa quorum-sensing regulator, NF-κB = Nuclear factor kappa-light-chain-enhancer of activated B cells, PI3K = Phosphoinositide 3-kinase, Akt = Protein kinase B, Nrf2 = Nuclear factor erythroid 2–related factor 2, MDA = Malondialdehyde, GFAP = Glial fibrillary acidic protein, Iba-1 = Ionized calcium-binding adapter molecule 1, ERK1/2 = Extracellular signal-regulated kinases 1/2, TNF-α = Tumor necrosis factor-alpha, GABA = Gamma-aminobutyric acid, BDNF = Brain-derived neurotrophic factor, JNK = c-Jun N-terminal kinase, MAPKs = Mitogen-activated protein kinases, PLpro = Papain-like protease, 3CLpro = 3-chymotrypsin-like protease, Main protease = SARS-CoV-2 main viral protease, ACE2 = Angiotensin-converting enzyme 2, TJP = Tight junction proteins, JAM-2 = Junctional adhesion molecule 2, MUC-2 = Mucin 2, HO-1 = Heme oxygenase-1, SOD = Superoxide dismutase, GSH = Reduced glutathione, GSSG = Oxidized glutathione, p53 = Tumor suppressor protein, PTEN = Phosphatase and tensin homolog, p21 = Cyclin-dependent kinase inhibitor, STAT3 = Signal transducer and activator of transcription 3, EGFR = Epidermal growth factor receptor, Wnt = Wingless-related integration site, P-gp = P-glycoprotein. Arrow line colour: red line=Quercetine, blue line=Kaempferol, green line=Apigenin, chocolateline line=Hesperetin, Yellow line=EGCG(Epigallocatechin Gallate), ↓= Inhibits or downregulates, ↑= upregulates.
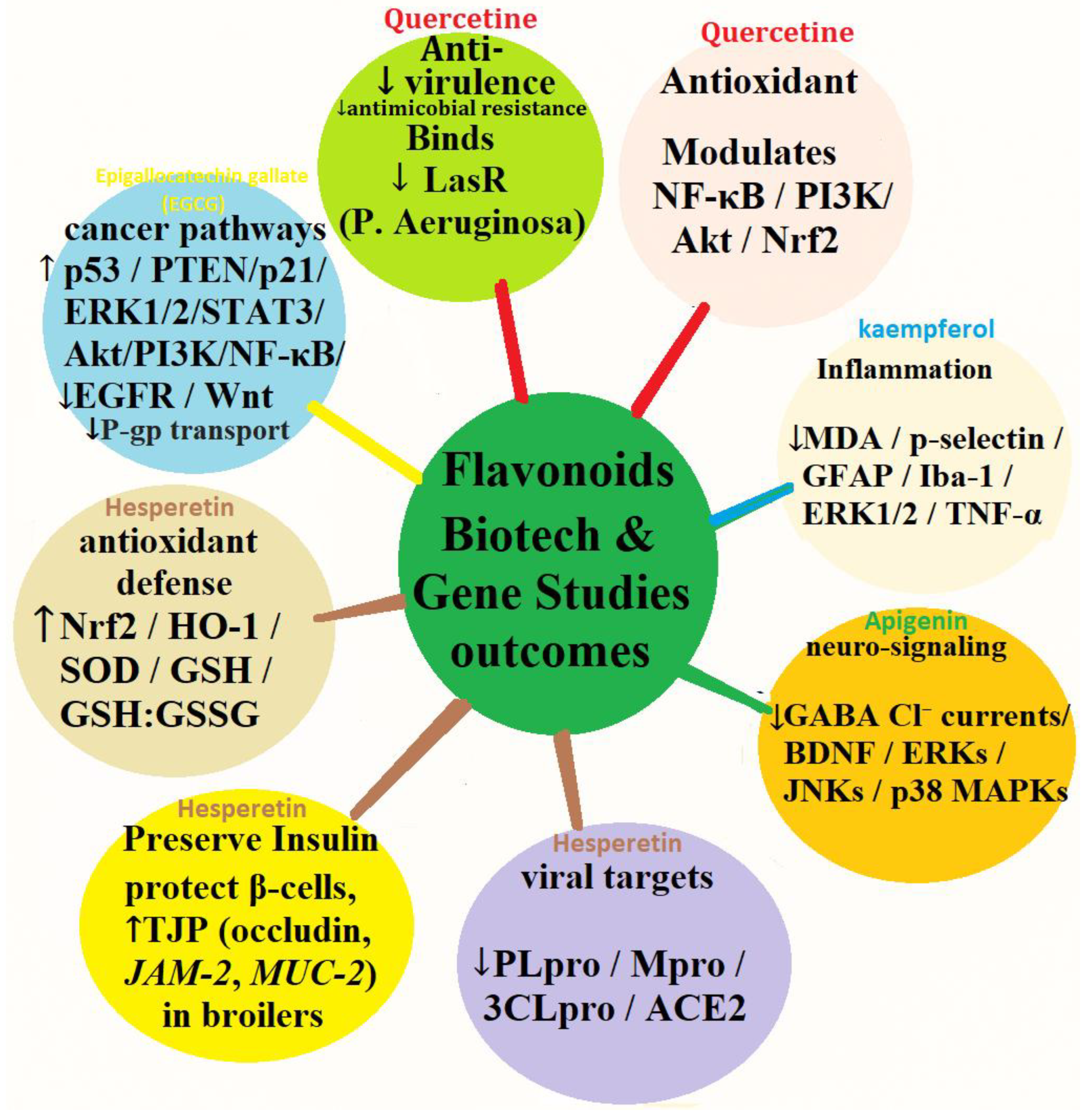
Figure 7.
Artemisinin-Mediated NRF2 Activation via KEAP1 Cysteine Modification (Modified from Pillai et al., 2022).
Figure 7.
Artemisinin-Mediated NRF2 Activation via KEAP1 Cysteine Modification (Modified from Pillai et al., 2022).
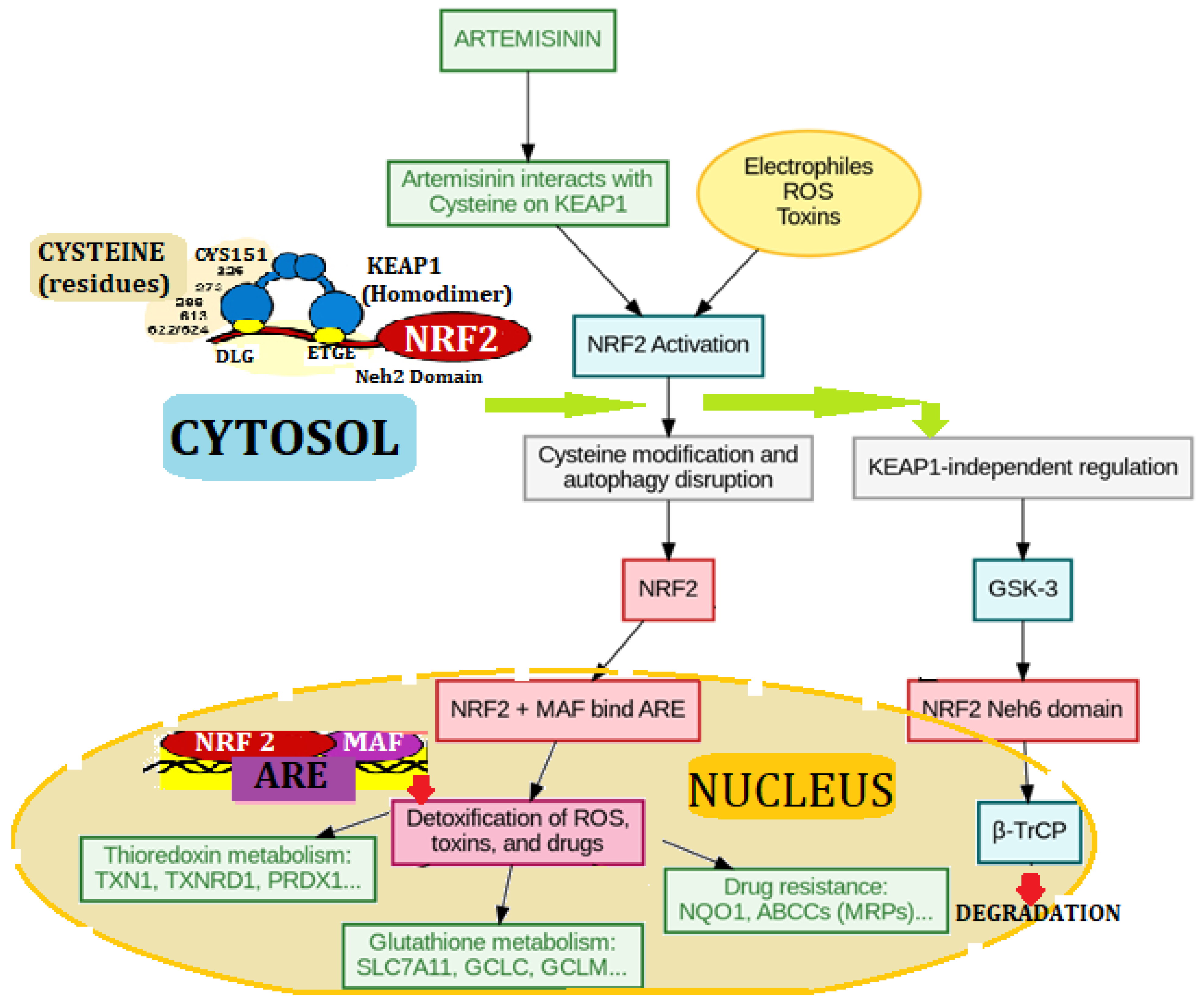

Table 1.
Comparative properties table of selected compounds classes: Sources, Properties and Biological Relevance.
Table 1.
Comparative properties table of selected compounds classes: Sources, Properties and Biological Relevance.
| Property / Compound | Gallic Acid | Quercetin | Not Available | Artemisinin and Solution |
|---|---|---|---|---|
| Chemical Class | Phenolics | Flavanols / Flavonoids (phenols) | Phenolics and unknown | Sesquiterpenes |
| Plant Sources | Phyllanthus emblica i(Amla) | Allium cepa (Onion) | Artemisia spp. roots | Artemisia annua |
| Antioxidant Assays | Free radical scavenging | DPPH, ABTS | DPPH | DPPH |
| DPPH Activity (%) | 68.53 | 79.8 | – | 91.0 ± 3.2 |
| MIC (mg/L) Against M.Os | 125 | 1000 | >256 | 14 |
| Enzyme Inhibition | Modifies hepatic drug metabolizing enzymes | Anti-cholinesterase activity | Inhibits α-amylase, α-glucosidase, tyrosinase, and cholinesterases | Acetylcholine esterase, digestive α-glucosidase |
| Biological Activities | Anti-tumor, antihepatotoxic, anti-inflammatory, antioxidant | Antioxidant, anticancer | Antioxidant, antimicrobial, anti-inflammatory, anti-Mycobacterium activity | Antimalarial, anti-inflammatory, antimicrobial, antibacterial, antioxidant |
| Absorption Route | Oral | Oral | Oral | Oral |
| Bioavailability | Low | Low | Low | Low |
| Toxicity (LD₅₀ mg/kg) | Rare, 5,000 in rabbits | 3807 | N.A. | >5,000 in rats |
| Solubility | Slightly soluble in water | Insoluble in water | Highly water-soluble | Sparingly soluble in water |
| Common Formulations | Nanosuspensions | Extract or waste-skin tablets | – | Artemisinin-containing solution or water extract and tablets |
| Research Status (2025) | Preclinical / clinical trials | Clinical | In vivo studies needed | For dental M.Os disease; clinical trials needed |
| Biotech & Gene Studies Outcomes | ↓ CYP450 enzyme activity, ↑ MAPKs, ↑ Nrf2, ↓ NF-κB, ↓ cytokines, ↓ Wnt/β-catenin, ↓ fungal protease | Binds LasR protein of P. aeruginosa, ↓ Violacein pigment, modulates NF-κB, PI3K/Akt, and Nrf2 | ↑ BZF metabolite, effects on COX-2 and TNF-α expression; antiproliferative (Rin-5F & HepG2 cells) | ↑ Keap1/Nrf2 pathway in sheep and humans |
| References | Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015 | Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020 | Trifan et al., 2022; Acquaviva et al., 2023 | Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025 |
Note: CYP450 = Cytochrome P450; MAPKs = Mitogen-Activated Protein Kinases; Nrf2 = Nuclear factor erythroid 2–related factor 2; NF-κB = Nuclear Factor kappa B; COX-2 = Cyclooxygenase-2; TNF-α = Tumor Necrosis Factor-alpha; Wnt/β-catenin = Cell signaling pathway; PI3K/Akt = Phosphoinositide 3-kinase/Protein Kinase B; HO-1 = Heme Oxygenase-1; Keap1 = Kelch-like ECH-associated protein 1; LasR = Quorum sensing regulator in P. aeruginosa; BZF = Benzofuran metabolite; Rin-5F = Rat insulinoma cell line; HepG2 = Human liver cancer cell line.
Table 2.
Comparative Properties Table of Selected Phenolic Acids: Properties and Biological Relevance.
Table 2.
Comparative Properties Table of Selected Phenolic Acids: Properties and Biological Relevance.
| Property / Compound | Gallic Acid | Caffeic Acid | Ellagic Acid | Syringic Acid | Salicylic Acid |
|---|---|---|---|---|---|
| Chemical Class | Phenolics, hydroxybenzoic | Phenolics/phenolic acidsm Hydroxycinnamic | Polyphenolic | Dimethoxybenzoic | Monohydroxybenzoic |
| Plant Sources | Phyllanthus emblica (Amla) |
Coffea arabica (Coffee) |
Punica granatum (Pomegranate) | Vitis vinifea (Grapes) | Salix alba Willow |
| Antioxidant Assays | DPPH, FRAP | DPPH, ABTS | DPPH, FRAP, LPO | DPPH, ABTS | DPPH, FRAP |
| DPPH Activity (%) | 68.53 | 93.4 | ~88 | 24.5 | 28.23(leaf) 48 (bark) |
| MIC (mg/L) Against M.Os | 125 | 1000 | 5-30 (H. pylori) | 625 | 500 |
| Enzyme Inhibition | Modifies hepatic drug metabolizing enzymes, ↓α-amylase |
inhibitor for low-density lipoprotein oxidative modification | ↓COX-2 & MAO-A enzymes | ↓NADPH oxidase, ↓iNOS, ↓COX-2, ↓Catalase, ↓ SOD, ↓ GPx |
↓AChE & BuChE, |
| Biological Activities | Anti-tumor, antihepatotoxic, anti-inflammatory, antioxidant | Antitumour, antioxidant, anti-inflammatory, anti cancer, antiaging, antiUVB damage, pathogenesis of atherosclerosis | Antioxidant, antidepressant, Antiapoptotic, anti-mutagenic, Antiviral, Hepatoprotective | antioxidant and anti-inflammatory, anticancer, ↓TNF-α, IL-6, IL-1β, IFN-γ | analgesic, antipyretic, antiinflammatory and antirheumatic, antimicrobial |
| Absorption Route | Oral | Oral | Oral | Oral | Absorbs readily in skin |
| Bioavailability | Low | Low | low | poor | poor |
| Toxicity (LD₅₀ mg/kg) | Rare, 5,000 in rabbits | 5000 in mice | >2000 in mice | >1,000 in rats | > 2000 in mice |
| Solubility | Slightly | Low | low | moderate | poor |
| Common Formulations | Nanosuspensions | encapsulation | Complex EA with cyclodextrins | polymer microcapsules | Aspirin, capsules |
| Research Status (2025) | Preclinical / clinical trials | In vivo | Pre-clinical | In vivo, preclinical, clinical trials needed | limited clinical studies |
| Biotech, molecular targets& Gene Studies Outcomes | ↓ CYP450 enzyme activity, ↑ MAPKs, ↑ Nrf2, ↓ NF-κB, ↓ cytokines, ↓ Wnt/β-catenin, ↓ fungal protease | ↓TAMs, ↓ NO ↓COX -2 and iNOS, ↓NF-κB, |
↑ Nrf2, ↓NF-κB, ↑cytoprotective genes—such as SOD, CAT, GPx, GR, GGT, GST, NQO1, and HO-1, ↓PGE₂, ↓TNF-α, ↓ IL-6, ↓IL-1β, ↓Bax/Bcl-2 ratio |
↑ KEAP1/NRF2, ↓ NF-κB, TLR4, HMGB1, MyD88, and TRAF6, ↑Ppara, ↑Cpt1, Cpt2, ↓ NF-κB, ↓ NRF1, ↓ MDA and ↓ TBARS, ↓CDK4&6, ↓Cyclins, ↓Bax, ↓Bak, ↓Bcl-2, ↓cIAP1&2, role in bioremediation | ↓ PGE2 Release, ↑NPR–TGA |
| References | Chen et al., 2025; Hasan et al., 2023; Singh and Kaur, 2015 | Khan et al., 2016; Alam et al., 2022b; Sousa et al., 2015; Faria et al., 2020; Mude et al., 2020 | Yang et al.,2023; Xin et al.,2017; Cervantes-Anaya et al., 2022; Evtyugin et al., 2020; verotta et al.,2018; De et al., 2018; Les et al., 2015; Pavlova et al., 2016; Naraki et al., 2022; Wojtunik-Kulesza et al.,2025 | Zhao et al., 2025; Srinivasulu et al., 2018; Rasheeed et al., 2025 | Le et al., 2021; Shalaby et al., 2025; Sultana et al., 2025;Zheng et al., 2024; Piątczak et al, 2020;Maistro et al.,2022; Randjelović et al., 2015; Jia et al, 2023; Chaudhary et al.,2023; Arif, 2015; Davidova et al ., 2024 |
Table 3.
Comparative properties table of Selected Flavonoids: Antioxidant and Biological Properties.
Table 3.
Comparative properties table of Selected Flavonoids: Antioxidant and Biological Properties.
| Property / Compound | Quercetin | Kaempferol | Apigenin | Hesperetin | EGCG |
|---|---|---|---|---|---|
| Chemical Class | Flavanols / Flavonoids (phenols) | Flavonol | Flavone | Flavanone | Flavonol |
| Plant Sources |
Allium cepa (Onion) |
Brassica oleracea (Kale) | Matricaria chamomilla, Chamomile, parsley | Citrus fruits (Orange) | Camellia sinensis (Green tea) |
| Antioxidant Assays | DPPH, ABTS | DPPH, ABTS |
DPPH | DPPH, ABTS | DPPH |
| DPPH Activity (%) | 79.8 | ~55 | 94.8 | 70 | 67.3 |
| MIC (mg/L) Against M.Os | 1000 | 256 | 2 | 62 & 500 | 200-400 |
|
Enzyme (inhibition) Studies |
Anti-cholinesterase activity | ↓AChE | ↓pancreatic lipase, ↓CD38 | ↓Mpro, PLpro, RdRp, & NSP15 |
↑caspases, ↓matrix metalloproteinases, ↓DHFR, ↓Telomerase, ↓MMPs in cancer cells |
| Biological Activities | Antioxidant, anticancer | antioxidant, antimicrobial, anticancer, neuroprotective, and hepatoprotective, Antigenotoxic, Antitumour | anti-inflammatory, antioxidant, analgesic, antimicrobial, hepatoprotective, anti-allergic, anticancer, and anti-hypertensive agent | Neuroprotective, antioxidative, antiviral, antidiabetic effects, preventing hepatic, renal and cerebral damage | anti-cancer, anti-oxidant, anti-inflammatory, anti-angiogenesis, anti apoptotic effects, anti bacterial |
| Absorption Route | Oral | Oral | Oral, GIT | orally & intraperitoneally | Oral, GIT |
| Bioavailability | Low | low | Low needs improvement | low | Low |
| Toxicity (LD₅₀ mg/kg) | 3807 | >2000 in rats | low | >4000 | > 45 in mice |
| Solubility | Insoluble | Low | diminished solubility | low | slight |
| Common Formulations | Extract or waste-skin tablets | nanoformulations | Nanoformulations, nanoemulsions, energy drinks | nanocomplexes | Extract, tea |
| Research Status (2025) | Clinical | In vivo, invitro | Animal, clinical | Clinical bioavailability improvement needed | Clinical |
| Biotech & Gene Studies Outcomes | Binds LasR protein of P. aeruginosa, ↓ Violacein pigment, modulates NF-κB, PI3K/Akt, and Nrf2 | ↓MDA, ↓p-selectin, GFAP, Iba-1, ERK1/2, & TNF-α, ↑Nrf2 |
↓ GABA (gamma-aminobutyric acid)-activated Cl− currents in rats, ↓BDNF, ↓ERKs, JNKs, and p38 MAPKs, | ↑Nrf2 & HO-1, protected RPE-19 cells from apoptosis, ↑SOD & GSH, ↑GSH/GSSG ratio, ↓PLpro , Mpro inhibition, ↓3CLpro main protease, Bind to ACE2, protect β-cells, ↑TJP (occludin, JAM-2, MUC-2) in broilers | ↑ p53 and PTEN/ p21, ↓ERK1/2, ↓ STAT3, ↓Akt/PI3K, NF-κB, ↓EGFR and ↓Wnt pathways , ↓DNA methylation in cancer, ↓P-gp transport |
| References | Quecan et al., 2019; Grzelak-Błaszczyk et al., 2018; Lee et al., 2015; Fuentes et al., 2020; Črnivec et al., 2021; Kandemir et al., 2024; Mishra et al., 2025; Dibal et al., 2020 | Bangar et al., 2023; Seema et al., 2023; Chandekar et al., 2022; Sharma et al., 2021; Hussain et al., 2022; Molaei et al., 2021; Jan et al., 2022 | Al-Dabbagh et al., 2019; Sah et al., 2022; Singh et al., 2024; Wang et al., 2019; Salehi et al., 2019; Siddiquee et al. 2025; Chen et al.¸2023a; Franco et al., 2020 | Khan et al., 2020; Tallei et al., 2020; Tomic et al., 2020; Agrawal et al., 2021; Wdowiak et al., 2022; Ciriminna et al., 2025; Zhao et al.,2023; Shrivastava et al., 2018; Choi et al., 2022 | Alam et al., 2022a; Cao et al.,2026; Amin et al.,2019; Nuryana et al.,2020; Furniturewall & Barve, 2022; Wang et al., 2015; Du et al., 2018 |
Table 4.
Comparative Antioxidant and Therapeutic Properties of Artemisinin and Its Derivatives .
| Property / Compound | Artemisinin | Semi-synthetic Artemisin derivatives (Dihydroartemisinin (DHA), Artesunate, Artemether, Arteether) |
|---|---|---|
| Chemical Class | Sesquiterpenes | Reduced artemisinin derivative |
| Plant/synthetic Sources | Artemisia annua | Metabolite of artemsinin, semi-syntheitic |
| Antioxidant Assays | DPPH, ABTS | N.A |
| DPPH Activity (%) | 91.0 ± 3.2 | N.A |
| IC₅₀ for DPPH (µg/mL) | 5.17 | N.A |
| MIC (mg/L) Against M.Os | 14 | N.A |
|
Enzyme (inhibition) Studies |
Acetylcholine esterase, digestive α-glucosidase, ↓PfATP6 | ↓PfATP6, modulates ↑ antioxidative enzymes |
| Biological Activities | Antimalarial, anti-inflammatory, antimicrobial, antibacterial, antioxidant, anti viral, antineoplastic activity against pancreatic, leukemic, osteosarcoma, and lung cancer cells | Anticancer, antitumor, anti-angiogenenic in cancer cells |
| Absorption Route | Oral | N.A |
| Bioavailability | Low | poor |
| Toxicity (LD₅₀ mg/kg) | >5,000 in rats | N.A |
| Toxicity | Neurotoxicity>28days in animals | N.A |
| Solubility | Sparingly soluble in water | Lipophilic, enhanced water solubility than artemisinin but still low |
| Common Formulations | Artemisinin-containing solution or water extract and tablets | |
| Research Status (2025) | For dental M.Os disease; clinical trials needed | Preclinical/clinical |
| Biotech, molecular targets & Gene Studies Outcomes | ↑ Keap1/Nrf2 pathway in sheep and humans, ↓IRE1α phosphorylation, Direct DNA damage to cancer, ↓ ERK1/2, ↓VEGFR2, ↑ antioxidative enzymes | ↑ ratio Bax/Bcl-2 ↑ caspase 3 & cytochrome c, ↓ AKT/GSK3β/cyclin D1 pathway, ↓TCTP, ↓HSP70, ↓Bcl-xL & Bcl-2, ↓AKT/SRC pathways inn breast cancer, ↓mTORC1, ↑ Nrf-2 and TRX |
| References | Kim et al., 2015; Siddiqui et al., 2018; Morua et al., 2025; Gang et al., 2025; Chen et al., 2024; Gavarić et al., 2025; Kuang et al.,2018; Wei and Liu, 2017; Moore et al., 2022 | Dai et al., 2021; Osonwa & Hu, 2018; Addissouky, 2025; Feng et al., 2016; Wei and Liu, 2017; Moore et al., 2022; Kazmi et al., 2024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



