
Submitted:
03 March 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
More than three decades have passed since adult T-cell leukemia/lymphoma (ATL) was first identified as a distinct disease in Japan. During this time, researchers discovered its causative agent—the human T-cell leukemia virus type 1 (HTLV-1). HTLV-1 infection has a significant global prevalence, affecting an estimated 5–10 million individuals worldwide. In this review, we provide a comprehensive overview of HTLV-1 persistence, latency, replication, and oncogenesis, with a particular focus on the roles of viral proteins such as Tax, HBZ, Rex, p30, p12, and p8. Understanding these mechanisms is essential for developing effective therapeutic strategies against HTLV-1-associated diseases, including ATL. Additionally, we explore current treatment strategies for ATL, including conventional chemotherapy, allogeneic hematopoietic cell transplantation, and epigenetic therapies. Gaining deeper insights into HTLV-1 pathogenesis, its molecular drivers and the existing therapeutic approaches can pave the way for more refined and innovative strategies to combat HTLV-1-associated malignancies.
Keywords:
ATL
; chemotherapy
; epigenetic therapy
; HBZ protein
; HTLV-1
; monoclonal antibodies
; stem cell transplantation
; tax protein
1. Introduction
The first identified human retrovirus, HTLV-1, has long been known to be linked to adult T-cell leukemia/lymphoma (ATL) and other serious illnesses [1]. HTLV-1 infection affects millions of people globally, with endemic regions comprising Gabon, the Democratic Republic of Congo, Nigeria, aboriginal communities in Australia, Equatorial Africa, Latin America, the Caribbean, northeastern Iran, and southwestern Japan [2]. Although most infected people do not exhibit any symptoms, 3-5% of infections will result in ATL or HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) decades after the initial infection [1]. HTLV-1 mainly targets CD4+ T lymphocytes and transforms them as a result of the expression of viral oncoproteins such as Transactivator protein X (Tax) and HTLV-1 basic leucine zipper (HBZ) [1]. ATL is a very aggressive form of leukemia with a dismal prognosis and few treatment options available [1]. Understanding the molecular mechanisms involved in HTLV-1 persistence, latency, and oncogenesis, has advanced significantly during the past few decades. However, and despite these explorations, ATL remains challenging to treat because the current medications can cause severe side effects and exhibit high relapse rates. Allogeneic hematopoietic cell transplantation and targeted therapies are examples of promising treatments that provide new hope for improving patient outcomes.
To provide a comprehensive understanding of HTLV-1-associated ATL, this review explores key topics and critical insights. It first examines the HTLV-1's biological makeup, including the virus's DNA, modes of transmission, and target cells. Furthermore, it examines the molecular mechanisms underlying viral replication and the persistence of HTLV-1 within host cells. The review also examines the mechanisms underlying HTLV-1-induced oncogenesis emphasizing the pivotal roles of key viral proteins such as Tax and HBZ in driving the transformation of immune cells. In addition, the study examines the current approaches to treating ATL, which include chemotherapy, targeted therapies, hematopoietic stem cell transplantation and epigenetic therapies. Lastly, the challenges encountered in treating HTLV-1-related ATL are discussed, including the disease's rarity and heterogeneity.
2. HTLV-1 Epidemiology
HTLV-1, the first human retrovirus ever identified, belongs to the Delta subtype of the Orthoretrovirinae family and is estimated to infect approximately 10 million people worldwide. [1]. High infection rates of this virus have been reported in Gabon and the Democratic Republic of Congo, while Nigeria has the largest number of infected individuals globally. Other endemic regions include aboriginal communities in Australia, Equatorial Africa, Latin America, the Caribbean, northeastern Iran, and southwestern Japan, with Romania and Moldova being the only European countries with relatively high seroprevalence [2]. This virus is associated with ATL, HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP), and various inflammatory conditions [1]. ATL, a particularly aggressive malignancy, develops in approximately 5% of infected individuals after 30 to 50 years, with a median survival time ranging from 6 to 24 months [1].
3. HTLV Genotype and Biological Structure
Although HTLV-1 is the most common and well-studied type of viruses, there are also other known genotypes named HTLV2/3 and 4 [3]. In addition, HTLV-1 is present in 7 subtypes (HTLV-1A to -1G) that are highly conserved [3] . Following a prolonged incubation period, only a small proportion of individuals infected with HTLV-1 progress to one of its associated diseases. These include ATL, an aggressive leukemia driven by the virus's oncogenic activity; HAM/TSP, a neurological demyelinating disorder; and HTLV-1-associated uveitis [4,5].
HTLV-1 is a single-stranded, positive-sense RNA virus. Its genome contains long terminal repeats (LTRs) at both ends [6], and it encodes the classical retrovirus structural proteins; group-specific antigen (gag), envelope (env) genes, polymerase (pol), reverse transcriptase (RT) and integrase (IN) [7]. Also, HTLV-1 encodes two oncoproteins, Tax and HBZ. Tax is responsible for cytoplasmic signaling through various receptors and leads to unregulated cell cycle, genetic instability, inhibition of DNA repair and finally apoptosis. While, HBZ is involved in inflammation, histone methylation, apoptosis and T-cell differentiation signaling pathways [7]. Moreover, HTLV-1 encodes for Regulator of Expression (Rex) a viral RNA-binding protein, that facilitates the transport of unspliced or incompletely spliced viral mRNAs from the nucleus to the cytoplasm of the host cell [6].
HTLV-2 is less pathogenic than HTLV-1 [3]. Interestingly, HTLV-2 has no clinical correlation with lymphoproliferative disease, even if it was first discovered in a patient with hairy cell leukemia [8]. In addition, some patients with HTLV-2 infection report HAM/TSP-like symptoms, but a clear correlation between the virus and symptoms needs to be established [9].
HTLV-3 and HTLV-4, two novel human T-cell leukemia viruses, were identified in Cameroon [3]. Their clinical significance and associated disorders remain under investigation due to limited research and understanding.
4. HTLV-1 Transmission and Infection
HTLV-1 was first detected in humans in Central Africa, originating from cross-species transmission of simian T-cell leukemia virus type-1 (STLV-1) from primates to humans. Three types of transmission were observed: human to human parallel transmission, primate to human transmission, and non-human transmission [3]. Additionally, the geographic movement of people helped in the spread of HTLV-1 in the world mainly via body fluid through three routes: sexual intercourse (semen), breastfeeding, and blood transfusion [6]. A Japanese study showed that the transmission from male to female is more efficient than from female to male [10]. Interestingly, the cell-free infection of HTLV-1 is inefficient [11]. HTLV-1's limited genetic variation suggests that it spreads by cell transmission rather than viral particle infection. Viruses propagate via virological synapses, biofilm, cellular conduits, and tunneling nanotubes [12] (Figure 1). Heparan sulfate proteoglycan (HSPG), neuropilin-1, and glucose transporter (GLUT1) are involved in HTLV-1 cell entrance. The virus should initially connect with HSPG, then neuropilin-1, and lastly GLUT-1 on the cell surface. This helps membrane fusion and viral entrance into cells [13].
5. Mechanisms of Viral Replication, Persistence and Oncogenesis
5.1. Replication Cycle: From Invasion to Mature Virions
While CD4+ T cells are the primary target of HTLV-1 infection, there is evidence that the virus also infects other types of immune cells, such as CD8+ T lymphocytes and B lymphocytes [3]. This is due to the ability of the envelope glycoprotein surface subunit to interact with three main cellular receptors including GLUT1, HSPG and the neuropilin-1 that are found widely throughout this range of target cells. The invasion of HTLV-1 represents the first step of the virus’s ability to survive and replicate. HTLV-1 envelope surface subunit initially interacts with HSPG followed with neuropilin-1 resulting in the creation of a complex. This association causes a conformational change in surface subunit triggering GLUT1 to join the complex for initiating the fusion process (Figure 2-1). Subsequently, the viral core containing the viral genomic RNA (gRNA) is delivered into the cytoplasm (Figure 2-2). Following its entry, the viral genomic RNA is reverse transcribed into a double stranded DNA (dsDNA) (Figure 2-3) and translocated into the nucleus (Figure 2-4). Afterward a provirus is formed by integration of viral genome into the host cell DNA (Figure 2-5 and 6). Consequently, the provirus is transcribed (Figure 2-7) and post transcriptionally modified (Figure 2-8) and then exported outside of the nucleus (Figure 2-9) followed by translation of the viral proteins in the cytoplasm (Figure 2-10). Two copies of the viral genomic RNA along with the structural and enzymatic proteins (Gag, pro, pol and env) are delivered to the plasma membrane (Figure 2-11) to assemble an immature virus budding (Figure 2-12) that is liberated from cell surface (Figure 2-13) and undergoes various maturation process to form a fully infectious virus particle (Figure 2-14) [14].
5.2. HTLV-1 Persistence and Latency: A Virus Hidden in a Plain Sight
In the early stage of infection and based on cellular host pathway, the doubly spliced mRNA Tax and Rex from HTLV-1 provirus are expressed to activate viral replication and expression of viral genes [15]. Tax stimulates the 5’LTR by binding to its Tax response element (TRE) via transcription Factor cAMP response element binding protein (CREB). This complex (Tax/CREB) called up cellular transcriptional co-activators to activate the 5’LTR viral promoter and enhance transcription of single spliced and un-spliced mRNA that codes for structural and enzymatic viral proteins [16,17]. Viral replication requires the nuclear export of these viral mRNA from site of transcription to the translation machinery in the cytoplasm. Rex the accessory protein of HTLV-1 plays a key role in this nuclear export. This leads to the accumulation of those unstable, not completely spliced mRNA in the cytoplasm and enhances their protein translation, which are required for replication of viral particle and virions formation [15]. After infecting T cells, HTLV-1 enters a prolonged phase of clinical latency in carriers, typically lasting between 20 and 40 years. During this time, viral gene expression remains minimal, making it difficult to detect and allowing the virus to evade immune surveillance [18]. Several accessory proteins, including p30, p12, and p8, play distinct roles in promoting viral latency and persistence through multiple mechanisms. p30 is a protein that resides in the nucleus and binds to doubly spliced mRNA, ensuring the retention of transcripts that encode the Tax and Rex proteins within the nucleus. This leads to a decrement in the protein levels and suppression of viral replication which indeed promotes persistence by evading the immune response [19]. On the other hand, p12 and p8 reduce the surface expression of intercellular cell adhesion molecule 1 (ICAM-1), ICAM-2, and Major Histocompatibility Class I (MHC-I), hindering the ability of NK cells and cytotoxic T lymphocytes to detect infected cells. Additionally, p8 suppresses early T-cell receptor signaling following T-cell receptor stimulation, thereby preventing T-cell activation [20]. Furthermore, given that Tax protein is strongly immunogenic, its expression should be inhibited to prevent the elimination of Tax expressing cells by Tax specific cytotoxic T lymphocytes. The repression of Tax is therefore controlled by different possible mechanisms including genetic changes in the Tax gene, epigenetic changes in the viral promoter in the 5’ LTR such as DNA hypermethylation and histone modifications, and/or deletion of 5’ LTR sequences [21]. It is important to note that the export of the mRNAs encoding HBZ occurs independently of Rex. Notably, HBZ can inhibit the Rex-mediated export of structural mRNAs (e.g., Gag-Pol), thereby preventing viral translation and preserving a latent HTLV-1 infection without viral production. Consequently, HBZ does not support productive viral replication, but rather contributes to HTLV-1 persistence by suppressing cellular senescence and promoting viral latency [22]. Together, these factors facilitate the establishment of a dormant state in HTLV-1-infected cells, allowing the virus to persist undetected within the host.
5.3. HTLV-1-Induced Cellular Oncogenesis: Insights into Key Viral Proteins and Their Regulatory Roles
ATL subtypes can be classified based on Tax expression. In about 50% of ATL cases, the expression of Tax is lost. Around 25% of cases originate from HTLV-1-infected cells that never expressed Tax, indicating Tax-independent leukemogenesis. Another 25% initially require Tax for transformation but later lose its expression to evade immune detection. The remaining 50% of ATL cases retain Tax expression, with two patterns: a small subset shows high Tax expression, while most cases express Tax intermittently or at very low levels [23].
Whole-genome sequencing analyses of ATL have highlighted two striking features of ATL that are connected to Tax: the extensive genomic instability observed in ATL, and the significant overlap between the signaling pathways and molecules activated by Tax and the genetic alterations found in ATL. These alterations predominantly affect key signaling molecules within the TCR-NF-κB and CD28 co-stimulatory pathways, underscoring Tax's role in disease progression [24].
Tax drives the transformation of infected T cells through multiple regulatory mechanisms. Tax disrupts cell cycle regulation by promoting the transcription of S-phase genes through Rb hyperphosphorylation, increasing WIP1 phosphatase activity, and suppression of CDK4 inhibitors, including p15, p16, p18, and p19, as well as the tumor suppressor p53 [25]. Additionally, Tax facilitates the transition into M phase by activating the anaphase-promoting complex (APC) and its co-activator cdc20. Furthermore, Tax induces centrosome amplification, contributing to aneuploidy and genomic instability [25]. On the other hand, Tax suppresses apoptosis by upregulating anti-apoptotic proteins, including X-linked inhibitor of apoptosis (XIAP), survivin, BCL-2, Bcl-xL, Bfl-1/A1, and cFLIP [25]. Additionally, it disrupts caspase signaling by inactivating key apoptotic effectors such as caspase-3, -7, -8, and -9, thereby promoting the survival of infected cells [25]. Additionally, Tax regulates miRNA expression to precisely modulate gene expression for cellular transformation. It enhances the proliferation of infected T cells by upregulating miR-146a, miR-130b, and miR-155 while downregulating miR-149, miR-135b, miR-872, and miR-873, thereby influencing key regulatory pathways involved in cell growth and survival [25].
In contrast to Tax, HBZ is expressed in all ATL cases [23] and is known to promote T-cell proliferation through multiple regulatory mechanisms [26]. First, HBZ was shown to activate the transcription of the survivin gene, a key inhibitor of apoptosis [26]. Second, HBZ protein was suggested to form heterodimers with C/EBPα imparing its DNA binding capacity and therefore overcoming its suppressive effect on cell proliferation [26]. Similarly, HBZ can form heterodimers with JunD leading to the activation of JunD-dependent cellular genes including human telomerase reverse transcriptase [26]. Moreover, HBZ modulates the Wnt signaling pathways, a process associated with the leukemogenesis of ATL. At the miRNA level, HBZ can also upregulate two oncogenic miRNAs (miR-17 and miR-21) at the post-transcriptional level [26]. In addition, HBZ-mediated miRNA regulation leads to OBFC2A downregulation, a process that drives both cell proliferation and genomic instability [26].
6. Therapeutic Strategies of HTLV-1 Associated Cancer
6.1. Watch-and-Wait Strategy for Indolent ATL
The choice of a therapeutic approach for ATL depends on the subtype of the disease. Indolent ATL subtypes, such as smoldering and favorable chronic, generally have a better short-term prognosis compared to aggressive forms. Traditionally, a "watch-and-wait" approach was used for these subtypes. However, a study in Brazil revealed that patients with indolent ATL who were managed with this approach had a 5-year survival rate of less than 20% [27]. Therefore, patients with slow-growing ATL should receive active treatment, as the watch-and-wait strategy is associated with poor long-term outcomes.
6.2. Chemotherapy
6.3. Immunomodulatory Therapy
Lenalidomide , an immunomodulator, demonstrated an antitumor activity and a manageable toxicity profile in patients with relapsed or recurrent aggressive ATL. Clinical outcomes included an overall response rate of 42% and a tumor control rate of 73%. The median progression-free survival and overall survival were 3.8 and 20.3 months, respectively. Adverse events, including neutropenia, leukopenia, lymphopenia, and thrombocytopenia, were generally manageable and reversible [34].
6.4. Antiviral Therapy
Zidovudine (AZT) is the only antiviral medicine utilized in the treatment of HTLV-1. Adoption of AZT and interferon-alfa (IFN-alpha) in ATL showed enhanced survival in the leukemic subtypes. The five-year overall survival rate was 46% for patients who received first-line antiviral therapy, 20% for patients who received first-line chemotherapy, and 12% for patients who received first-line chemotherapy followed by antiviral therapy. Patients with acute, chronic, and smoldering ATL significantly benefited from first-line antiviral therapy, whereas patients with ATL lymphoma experienced a better outcome with chemotherapy. In acute ATL, achievement of complete remission with antiviral therapy resulted in 82% 5-year survival. Antiviral therapy in chronic and smoldering ATL resulted in 100% 5-year survival [35]. Mechanistically, AZT/IFN-alpha reduces the reverse transcriptase activity and alters the clonality pattern in ATL patients [36]. However, although there was clinical improvement, the treatment with AZT/IFN did not cure patients with acute and lymphoma ATL, who still had medical needs that were not being satisfied [37].
6.5. Arsenic Trioxide (As2O3)- Based Therapy
The combination of As2O3 and INF-alpha selectively eliminated ATL cells by promoting Tax proteasomal degradation and inhibited the activity of ATL leukemia initiating cells in vivo [38,39,40]. At the molecular level, the exact mechanism of action of As2O3 is not yet fully understood; however, it seems to promote apoptosis in treated cells. A strong synergistic impact was noted between IFN-alpha and arsenic in ATL-derived cell lines, leading to a significant suppression of cell proliferation, G1 arrest, and induction of apoptosis [38]. Cell death was shown to be partially reliant on caspase activation and was not linked to changes in bcl-2, bax, or p53 expression. As2O3 and IFN-alpha together caused Tax degradation, upregulation of IkB-a leading to a reduction in RelA DNA binding nuclear factor NF-kB complexes due to the cytoplasmic retention of RelA. Assuming the function of Tax in HTLV-I–induced transformation, its decrease likely leads to cell death by deactivating the NF-kB pathway [41].
In clinical settings, a combination regimen of As2O3, IFN-alpha, and AZT was evaluated in 10 newly diagnosed patients with chronic ATL, achieving a remarkable 100% response rate. This included seven complete remissions, two complete remissions with residual circulating atypical lymphocytes exceeding 5%, and one partial response. Notably, responses were rapid, adverse effects were moderate and primarily hematologic, and no relapses were observed [42].
Moreover, the combination of As2O3, IFN-alpha and AZT over 8 weeks showed potential as a consolidation therapy for ATL patients. Among nine patients, one patient in progression died. While three survivors with the indolent subtype achieved survival times of 48, 53, and 97 months and response durations of 22, 25, and 73 months. The five remaining patients had aggressive ATL. The median survival time was 36 months, and the median response duration was 10 months. The resulting side effects were manageable and reversible, reinforcing the feasibility of this treatment as a viable option for patients who are ineligible for allogeneic stem cell transplantation. [43].
Furthermore, the combination of As2O3, INF-alpha and AZT in a 30-day study involving 16 ATL patients demonstrated a shift from a Treg/Th2 to a Th1 cytokine profile, marked by increased IFN-γ and IL-2 levels and decreased Foxp3, IL-10, and IL-4 levels. Fifteen patients responded to treatment, showing reduced CD4+CD25+ cells and Foxp3 transcripts. The therapy improved the immune microenvironment, aiding ATL cell eradication and reducing opportunistic infection risks. The transcript levels of IL-10, an immunosuppressive cytokine upregulated by Tax, negatively correlated with clinical response, highlighting their significance in predicting treatment outcomes [44].
6.6. Monoclonal Antibody Therapy
Various monoclonal antibodies targeting cell surface molecules on ATL cells were evaluated in clinical studies with promising results. The monoclonal antibodies primarily targeted CD2, CD25, CD30, CD52, and TfR in ATL cells.
6.6.1. mAb A24 Targeting TfR
HTLV-1-infected cells constitutively express high levels of surface transferrin receptor (TfR), a feature not observed in normal cells. The anti-TfR antibody A24 was shown to inhibit [55Fe]-transferrin uptake in activated T cells, downregulate TfR expression, and disrupt TfR recycling. Notably, A24 effectively suppressed the ex vivo proliferation of malignant T cells from both acute and chronic ATL by inducing programmed cell death, highlighting its potential as a therapeutic agent [45].
6.6.2. Alemtuzumab Targeting CD52
Alemtuzumab is a chimeric humanized antibody that was developed to target CD52 glycoprotein on ATL cells. Alemtuzumab was administered to 29 patients with ATL, achieving a response in 15 patients. While the treatment exhibited an acceptable toxicity profile, responses were of limited duration. The median progression-free survival was 2.0 months, and the median overall survival was 5.9 months. The most common adverse events included vasovagal episodes (7%), hypotensive episodes (10%), and hematologic toxicities such as leukopenia (41% Grade 3, 17% Grade 4), lymphocytopenia (59% Grade 3), neutropenia (31% Grade 3), anemia (24%), and thrombocytopenia (10%) [46].
6.6.3. MEDI-507 and an HAT Monoclonal Antibody Targeting CD2 and CD25 (IL-2 Receptor Alpha), Respectively
The evaluation of MEDI-507, a humanized monoclonal antibody targeting CD2, was conducted both independently and in combination with HAT targeting CD25 (IL-2 receptor alpha), using a human ATL xenograft model. Mice treated with MEDI-507 had a longer survival time compared to those treated with HAT. The results showed that MEDI-507 is effective in treating ATL and suggested that a clinical trial with this monoclonal antibody could be beneficial for patients with CD2-expressing leukemias and lymphomas [47]. On the other hand, treatment with HAT monoclonal antibody showed extended remission durations in ATL, though significant hematological toxicity was observed. Intravenous HAT administration led to mixed, partial, or complete remissions lasting from 9 weeks to over 3 years, accompanied by restored serum calcium levels, improved liver function, and sometimes enhanced immune status. Targeting the IL-2 receptor on ATL cells with monoclonal antibodies presents a promising therapeutic strategy for this aggressive cancer [48].
6.6.4. Brentuximab Vedotin Targeting CD30
A 63-year-old patient with severe ATL exhibiting strong CD30 expression achieved sustained remission after receiving an intensive treatment regimen incorporating Brentuximab Vedotin-based chemotherapy, followed by Brentuximab Vedotin maintenance therapy [49].
6.7. Allogeneic Hematopoietic Cell Transplantation
Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is being more commonly utilized as a curative treatment for ATL. A comprehensive systematic review and meta-analysis was conducted to extract and synthesize data on clinical outcomes from 18 studies evaluating the efficacy and risks of allo-HSCT in patients with HTLV-1-associated ATL. Despite the considerable heterogeneity among the included studies, the pooled post-allografting complete response rate was 73%, while overall survival and progression-free survival rates were 40% and 37%, respectively. Additionally, the pooled relapse and non-relapse mortality rates were 36% and 29%, respectively. This indicates that allo-HSCT is an effective treatment for patients with ATL. However, addressing and mitigating relapses remains crucial for achieving long-term disease control [50]. In 2005, an analysis of 40 allo-HSCT for acute and lymphoma-type ATL in Japan, using stem cells derived from bone marrow or peripheral blood, found a 3-year overall survival rate of 45.3%, with relapse-free survival at 33.8% and disease relapse at 39.3%. While all evaluable patients achieved complete remission, the median survival was 9.6 months. Among relapsed cases, five patients attained complete remission again, three of whom responded to reduced or discontinued immunosuppressive therapy, suggesting a graft-versus-ATL effect. However, graft-versus-host disease did not appear to influence relapse prevention. These findings support allo-HSCT as a potential treatment option for select patients with aggressive ATL [51]. In 2010, a study analyzing 386 ATL patients undergoing allo-HSCT with different graft sources found a 3-year overall survival rate of 33%. Multivariable analysis identified four factors associated with lower survival: age over 50, male sex, non-remission status at transplantation, and the use of unrelated cord blood compared to HLA-matched related grafts. Treatment-related mortality was higher among cord blood recipients, while disease-associated mortality was elevated in male patients and those transplanted while not in remission. Additionally, in related transplants, donor HTLV-I seropositivity was linked to worse disease-associated mortality, emphasizing the importance of donor selection and achieving remission prior to transplantation for improved outcomes [52]. However, in 2012, a study of 10 patients with ATL undergoing unrelated cord blood transplantation found a 2-year overall survival rate of 40%. Prolonged survival was observed in chemosensitive patients, with four surviving beyond 500 days, while none of the chemorefractory patients survived. There was no engraftment failure, and acute graft-versus-host disease developed in seven patients, with grade 2 in three patients and grade 3 in four patients. These findings suggest that unrelated cord blood transplantation is a feasible treatment option, particularly for patients with chemosensitive ATL [53]. In 2014, a retrospective analysis of 175 patients with ATL who underwent single-unit cord blood transplantation reported a 2-year overall survival rate of 20.6%. Multivariate analysis identified the occurrence of graft-versus-host disease as a favorable prognostic factor for survival. However, the cumulative incidence of treatment-related mortality was high (46.1%), with early death being a significant concern. [54]. In 2020, a retrospective analysis of eight Romanian patients with aggressive ATL who received allo-HSCT showed a median post-transplant survival of 19.5 months. Four patients (50%) relapsed at a median of 5 months post-transplant. Graft-versus-host disease developed in six patients, and two patients died from Graft-versus-host disease -related complications. Despite high relapse rates, allo-HSCT was suggested to improve outcomes for aggressive ATL [55]. On the other hand, the administration Mogamulizumab (an antibody targeting CC chemokine receptor type 4) before allo-HSCT has been associated with an increased risk of severe acute graft-versus-host disease due to the reduction of donor-derived regulatory T cells [56]. However, a case report performed in 2020 describes an ATL patient who underwent allo-HSCT after Mogamulizumab treatment without developing severe graft-versus-host disease. The patient’s clinical course suggests that monitoring residual Mogamulizumab levels may help determine the optimal timing for allo-HSCT, potentially mitigating Mogamulizumab’s adverse effects on post-transplant outcomes without increasing the risk of relapse or progression [56].
In terms of side effects of allo-HSCT, most frequent cause of death was due to relapse. In addition, a 45-year-old patient developed unilateral panuveitis 18 months after undergoing allo-HSCT for lymphoma-type ATL. The inflammation resembled HTLV-1-associated uveitis but was caused by donor-derived CD8+ T cells, as confirmed by vitrectomy and fluorescence in situ hybridization. This suggests that donor T cells can induce panuveitis similar to HTLV-1-associated uveitis in ATL patients post-transplantation. The condition was recurrent but responded well to regional corticosteroid treatment [57].
Collectively, Allo-HSCT remains a viable and potentially curative option for ATL patients, but its long-term success is limited by high relapse rates and transplant-related complications. Optimizing donor selection, pre-transplant disease status, post-transplant monitoring, and immunosuppressive strategies will be critical in improving patient outcomes.
6.8. Targeting the Epigenetic Machinery
The Polycomb Repressive Complex 2 (PRC2) is a critical epigenetic regulator involved in gene silencing and transcriptional repression. Its function is mediated through the di- and trimethylation of lysine 27 on histone H3. This enzymatic activity is primarily driven by the EZH1 and EZH2 subunits, which serve as the catalytic core of PRC2. While EZH1 is expressed in both proliferating and differentiated cells, EZH2 is predominantly active in actively dividing cells [58].
Interestingly, integrative analyses of the epigenome and transcriptome in primary ATL cells and corresponding normal CD4⁺ T cells identified PRC2-mediated epigenetic reprogramming as a central mechanism underlying ATL pathogenesis. This reprogramming is suggested to occur at an early stage of ATL development, potentially in association with HTLV-1 infection. A global increase in the levels of trimethylation of lysine 27 on histone H3 was found to drive the ATL-specific gene expression landscape. This included the silencing of key tumor suppressor genes, transcription factors, epigenetic regulators, and microRNAs [59].
An in vitro study showed that ATL cells exhibited significantly higher EZH2 transcript levels, accompanied by enhanced trimethylation of lysine 27 on histone H3, compared to normal CD4⁺ T cells. Notably, patients with high EZH2 expression have a significantly worse prognosis, suggesting a potential role for EZH2 in ATL oncogenesis and disease progression. Pharmacological inhibition of histone methylation using 3-deazaneplanocin A effectively suppressed ATL cell proliferation. Furthermore, a synergistic effect was observed when 3-deazaneplanocin A was combined with the histone deacetylase inhibitor Panobinostat, highlighting a promising epigenetic-based therapeutic strategy for ATL treatment [60].
In the clinical setting, the efficacy and safety of valemetostat, a potent dual EZH2/EZH1 inhibitor, were evaluated for the treatment of relapsed or refractory ATL. A total of 25 patients (median 3 prior lines of therapy), including 24 previously treated with mogamulizumab, were enrolled. The centrally reviewed overall response rate was 48.0%, comprising 5 complete and 7 partial remissions. Treatment-emergent adverse events were manageable, supporting valemetostat’s potential as a promising therapeutic option for heavily pretreated ATL patients [61]. Notably, valemetostat became the first dual EZH1/EZH2 inhibitor approved for the treatment of aggressive ATL in Japan in September 2022 (reviewed in [62]).
6.9. Vaccines
Over the past decades, intensified efforts have been devoted to developing both traditional and advanced vaccine approaches for the therapeutic and preventive management of ATL.
In 1987, Nakamura et al. expressed HTLV-1 envelope gene products in Escherichia coli and used them to immunize cynomolgus monkeys. This approach elicited the production of antibodies against gp68 and gp46. Following viral challenge, no HTLV-1 antigens were detected in cultures of the immunized animals' peripheral blood mononuclear cells, suggesting that they were protected from HTLV-1 infection. [63]. In 1995, a recombinant vaccinia virus expressing the entire HTLV-1 envelope gene was assessed in rabbits. Although immunization induced anti-HTLV-1 envelope antibodies, it failed to generate neutralizing antibodies capable of preventing infection providing only temporary protection in one of three rabbits [64]. Another study conducted in the same year investigated an attenuated poxvirus expressing the full-length HTLV-1 envelope protein in New Zealand white rabbits. Initially, the vaccine conferred protection against HTLV-1 infection, but this immunity waned over time. All the previously protected rabbits became infected upon re-challenge five months later with HTLV-1-infected blood [65]. In 1997, a recombinant vaccinia virus expressing the HTLV-1 envelope gp46 protein under the control of a strong promoter induced the production of neutralizing antibodies and conferred a long-term protection against HTLV-1 infection in immunized cynomolgus monkeys. [66]. In 2000, Armand et al. explored DNA immunization strategies by testing plasmids encoding the HTLV-I envelope glycoprotein gene under different promoters in mice. The authors found that the construct driven by the human desmin muscle-specific promoter elicited enhanced humoral immune response and neutralization capacity, compared to constructs under the CMV promoter. These findings highlight the key role of vector design in optimizing the efficacy of the genetic vaccine against HTLV-I [67]. In an attempt to improve vaccine efficacy, researchers investigated the incorporation of multiple immunogenic epitopes. In 2006, a chimeric peptide vaccine composed of a B-cell epitope from the HTLV-1 envelope region (aa 175–218) and three HLA-A*0201-restricted cytotoxic T-lymphocyte epitopes from the Tax protein (Tri-Tax) was evaluated for immunogenicity and protective efficacy using a squirrel monkey model. Immunization induced high-titer antibodies and an elevated frequency of IFN-γ-producing cells specific to the envelope and Tri-Tax immunogens, whereas individual Tax peptides failed to elicit a comparable response. Interestingly, following viral challenge, immunized animals exhibited lower proviral loads than controls, indicating the induction of partial protection against HTLV-1 [68]. Later on, vaccine strategies evolved to incorporate more sophisticated immunization techniques, including dendritic cell -based therapies. In 2015, a vaccine comprising autologous dendritic cells pulsed with Tax peptides was developed and administered to three previously treated individuals in an attempt to amplify Tax-specific cytotoxic T lymphocyte responses in ATL patients. This immunotherapy successfully stimulated Tax-specific cytotoxic T lymphocyte responses, leading to an improvement in performance status without inducing severe side effects. Among the patients, two achieved partial remission, with one eventually attaining complete remission. The third patient maintained stable disease for the first eight weeks but later exhibited slow disease progression, potentially due to the lack of Tax expression in his tumor cells at the time of biopsy [69]. Another study conducted in the same year showed that a recombinant vaccinia virus expressing HBZ protein can induce protective immunity in mice. Notably, the transfer of CD4+ T cells from HBZ-transgenic mice into vaccinated recipients resulted in a significant reduction in the presence of these CD4+ T cells within the spleen, mesenteric, and inguinal lymph nodes. Additionally, the transfer of splenocytes from immunized mice to lymphoma cell-inoculated mice prolonged their survival and provided partial protection against the disease. In the light of these findings, the peptide sequence responsible for conferring this immune protection was identified to be HBZ157-176 making it a promising candidate for the development of HBZ peptide vaccine [70]. In 2023, a novel recombinant Modified Vaccinia Ankara and a plasmid DNA vaccine, both expressing a multiepitope protein incorporating HBZ-derived peptides (HBZ42–50 and HBZ157–176), were developed and tested using a prime-boost heterologous protocol. This approach, designed to leverage the unique advantages of each vector, resulted in a robust immune response characterized by the activation of cytotoxic T lymphocytes and the induction of polyfunctional T-cell responses, including IFN-γ, TNF-α, and IL-2 production. These findings introduced this candidate as a promising therapeutic vaccine for HTLV-1-associated diseases [71]. Most recently, in 2024, Tu et al. developed a codon-optimized HTLV-1 envelope (gp62) mRNA, delivered via lipid nanoparticles and assessed its immunogenicity and effectiveness against HTLV-1 infection in a rabbit model. Following the first viral challenge, half of the vaccinated rabbits (3 out of 6) were completely protected. After a second viral challenge, two of the previously protected rabbits sustained sterilizing immunity. This vaccine not only reduced viral load but also enhanced immune responses, including CD4+/IFN-γ+ and CD8+/IFN-γ+ cells. Furthermore, it induced detectable envelope antibodies and neutralizing activity, which correlated with the observed protection against the virus [72].
7. Challenges and Limitations in ATL Treatment
One of the primary challenges in HTLV related ALT treatment lies in the rarity of the disease itself [61]. HTLV-1-associated ATL is relatively uncommon compared to other forms of leukemia and lymphoma, making it difficult to conduct large-scale clinical trials to evaluate treatment efficacy. This scarcity of patients limits the available data and hampers the development of evidence-based treatment protocols. The heterogeneous nature of the clinical course of ATL presents a significant obstacle. The disease manifests in various clinical forms, ranging from acute, lymphoma, chronic, and smoldering types [73]. Each one may require a tailored approach to treatment, further complicating therapeutic development. Another critical challenge stems from the intricate relationship between HTLV-1 infection and the development of ATL. Unlike many other cancers, ATL arises from a pre-existing viral infection, necessitating a dual-targeted therapeutic approach that addresses both the cancerous cells and the underlying viral infection. This multifaceted nature of the disease demands innovative treatment strategies that target HTLV-1-infected cells while sparing healthy tissues. Furthermore, the limited understanding of HTLV-1 pathogenesis poses a significant barrier to treatment development. Although researchers have made significant strides in unraveling the mechanisms underlying HTLV-1 infection and its association with ATL, many aspects of the disease remain elusive. The lack of comprehensive knowledge hinders the identification of druggable targets and the development of targeted therapies.
8. Conclusion
HTLV is a complex retrovirus that infects T cells and can lead to various diseases, including immunological, neurological disorders, and cancer. There are four known genotypes of HTLV, with HTLV-1 being the most common and well-studied type. The virus enters cells through interactions with specific molecules on the cell surface, primarily targeting CD4+ T cells but can also infect other immune cells. HTLV-1 infection leads to viral persistence in host cells through the activation of viral replication by proteins like Tax and Rex, followed by a shift into a deep latency period lasting for decades. The oncogenic transformation of infected CD4+ T cells is regulated by the key viral proteins Tax and HBZ. Treatment strategies for HTLV-1 associated cancer are numerous. While all the treatment strategies showed promising results, none can be considered a truly curative approach at this time. Chemotherapy alone has limited effectiveness in ensuring the long-term survival of ATL patients, highlighting the need for exploring new treatment approaches. The combination of As2O3, INF-alpha and AZT showed promising response rates with manageable side effects. Allo-HSCT is a potentially curative treatment for ATL. However, the number of patients who can undergo transplantation is limited, and the leading cause of mortality post-transplantation is relapse and transplant toxicity. Efforts to develop safe and effective HTLV-1 vaccinations are ongoing, with research focusing on various proteins and peptides associated with the virus. mRNA-based vaccines show promise in preventing HTLV-1 infection and persistence, with potential to improve immune responses and reduce viral loads. Challenges in treating HTLV-related ATL include the rarity of the disease, heterogeneous clinical forms, dual-targeted therapeutic approaches, and limited understanding of pathogenesis. Innovative treatment strategies are needed to address the multifaceted nature of HTLV-1 infection and ATL, considering the complex relationship between the virus and cancer development.
Funding
This review was not supported by any funds.
Declaration on interests
The authors declare that they have no conflict of interest.
Acknowledgments
We thank Miss Zaynab Fatfat for helping in the literature search and editing the final manuscript.
References
- Marino-Merlo, F., Grelli, S., Mastino, A., Lai, M., Ferrari, P., Nicolini, A., Pistello, M., & Macchi, B. (2023). Human T-Cell Leukemia Virus Type 1 Oncogenesis between Active Expression and Latency: A Possible Source for the Development of Therapeutic Targets. International journal of molecular sciences, 24(19), 14807. [CrossRef]
- Gessain, A., & Cassar, O. (2012). Epidemiological Aspects and World Distribution of HTLV-1 Infection. Frontiers in microbiology, 3, 388. [CrossRef]
- Zhang, L. L., Wei, J. Y., Wang, L., Huang, S. L., & Chen, J. L. (2017). Human T-cell lymphotropic virus type 1 and its oncogenesis. Acta pharmacologica Sinica, 38(8), 1093–1103. [CrossRef]
- Saito M. (2019). Association Between HTLV-1 Genotypes and Risk of HAM/TSP. Frontiers in microbiology, 10, 1101. [CrossRef]
- Kamoi, K., Watanabe, T., Uchimaru, K., Okayama, A., Kato, S., Kawamata, T., Kurozumi-Karube, H., Horiguchi, N., Zong, Y., Yamano, Y., Hamaguchi, I., Nannya, Y., Tojo, A., & Ohno-Matsui, K. (2022). Updates on HTLV-1 Uveitis. Viruses, 14(4), 794. [CrossRef]
- Nakano, K. , Yokoyama, K., Shin, S., Uchida, K., Tsuji, K., Tanaka, M., Uchimaru, K., & Watanabe, T. (2022). Exploring New Functional Aspects of HTLV-1 RNA-Binding Protein Rex: How Does Rex Control Viral Replication?. Viruses, 14(2), 407. [CrossRef]
- Ratner L. (2022). A role for an HTLV-1 vaccine?. Frontiers in immunology, 13, 953650. [CrossRef]
- Kalyanaraman, V. S., Sarngadharan, M. G., Robert-Guroff, M., Miyoshi, I., Golde, D., & Gallo, R. C. (1982). A new subtype of human T-cell leukemia virus (HTLV-II) associated with a T-cell variant of hairy cell leukemia. Science (New York, N.Y.), 218(4572), 571–573. [CrossRef]
- Martinez, M. P., Al-Saleem, J., & Green, P. L. (2019). Comparative virology of HTLV-1 and HTLV-2. Retrovirology, 16(1), 21. [CrossRef]
- Kajiyama, W., Kashiwagi, S., Ikematsu, H., Hayashi, J., Nomura, H., Okochi, K., Intrafamilial Transmission of Adult T Cell Leukemia Virus, The Journal of Infectious Diseases, 154(5), 851–857. [CrossRef]
- Derse, D., Hill, S. A., Lloyd, P. A., Chung Hk, & Morse, B. A. (2001). Examining human T-lymphotropic virus type 1 infection and replication by cell-free infection with recombinant virus vectors. Journal of virology, 75(18), 8461–8468. [CrossRef]
- Al Sharif, S., Pinto, D. O., Mensah, G. A., Dehbandi, F., Khatkar, P., Kim, Y., Branscome, H., & Kashanchi, F. (2020). Extracellular Vesicles in HTLV-1 Communication: The Story of an Invisible Messenger. Viruses, 12(12), 1422. [CrossRef]
- Yasunaga, J., & Matsuoka, M. (2011). Molecular mechanisms of HTLV-1 infection and pathogenesis. International journal of hematology, 94(5), 435–442. [CrossRef]
- Martin, J. L., Maldonado, J. O., Mueller, J. D., Zhang, W., & Mansky, L. M. (2016). Molecular Studies of HTLV-1 Replication: An Update. Viruses, 8(2), 31. [CrossRef]
- Nakano, K., & Watanabe, T. (2022). Tuning Rex rules HTLV-1 pathogenesis. Frontiers in immunology, 13, 959962. [CrossRef]
- Kashanchi, F., & Brady, J. N. (2005). Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene, 24(39), 5938–5951. [CrossRef]
- Boxus, M., Twizere, J. C., Legros, S., Dewulf, J. F., Kettmann, R., & Willems, L. (2008). The HTLV-1 Tax interactome. Retrovirology, 5, 76. [CrossRef]
- Kannian, P., & Green, P. L. (2010). Human T Lymphotropic Virus Type 1 (HTLV-1): Molecular Biology and Oncogenesis. Viruses, 2(9), 2037–2077. [CrossRef]
- Nicot, C., Dundr, M., Johnson, J. M., Fullen, J. R., Alonzo, N., Fukumoto, R., Princler, G. L., Derse, D., Misteli, T., & Franchini, G. (2004). HTLV-1-encoded p30II is a post-transcriptional negative regulator of viral replication. Nature medicine, 10(2), 197–201. [CrossRef]
- Edwards, D., Fenizia, C., Gold, H., de Castro-Amarante, M. F., Buchmann, C., Pise-Masison, C. A., & Franchini, G. (2011). Orf-I and orf-II-encoded proteins in HTLV-1 infection and persistence. Viruses, 3(6), 861–885. [CrossRef]
- Zane, L., & Jeang, K. T. (2014). HTLV-1 and leukemogenesis: virus-cell interactions in the development of adult T-cell leukemia. Recent results in cancer research. Fortschritte der Krebsforschung. Progres dans les recherches sur le cancer, 193, 191–210. [CrossRef]
- Ma, G. Ma, G., Yasunaga, J., & Matsuoka, M. (2016). Multifaceted functions and roles of HBZ in HTLV-1 pathogenesis. Retrovirology, 13, 16. [CrossRef]
- Nosaka, K., & Matsuoka, M. (2021). Adult T-cell leukemia-lymphoma as a viral disease: Subtypes based on viral aspects. Cancer science, 112(5), 1688–1694. [CrossRef]
- Giam, C. Z., & Semmes, O. J. (2016). HTLV-1 Infection and Adult T-Cell Leukemia/Lymphoma-A Tale of Two Proteins: Tax and HBZ. Viruses, 8(6), 161. [CrossRef]
- Mohanty, S., & Harhaj, E. W. (2020). Mechanisms of Oncogenesis by HTLV-1 Tax. Pathogens (Basel, Switzerland), 9(7), 543. [CrossRef]
- Zhao T. (2016). The Role of HBZ in HTLV-1-Induced Oncogenesis. Viruses, 8(2), 34. [CrossRef]
- Bittencourt, A. L., da Graças Vieira, M., Brites, C. R., Farre, L., & Barbosa, H. S. (2007). Adult T-cell leukemia/lymphoma in Bahia, Brazil: analysis of prognostic factors in a group of 70 patients. American journal of clinical pathology, 128(5), 875–882. [CrossRef]
- Taguchi, H., Kinoshita, K. I., Takatsuki, K., Tomonaga, M., Araki, K., Arima, N., Ikeda, S., Uozumi, K., Kohno, H., Kawano, F., Kikuchi, H., Takahashi, H., Tamura, K., Chiyoda, S., Tsuda, H., Nishimura, H., Hosokawa, T., Matsuzaki, H., Momita, S., Yamada, O., … Miyoshi, I. (1996). An intensive chemotherapy of adult T-cell leukemia/lymphoma: CHOP followed by etoposide, vindesine, ranimustine, and mitoxantrone with granulocyte colony-stimulating factor support. Journal of acquired immune deficiency syndromes and human retrovirology : official publication of the International Retrovirology Association, 12(2), 182–186. [CrossRef]
- Yamada, Y., Tomonaga, M., Fukuda, H., Hanada, S., Utsunomiya, A., Tara, M., Sano, M., Ikeda, S., Takatsuki, K., Kozuru, M., Araki, K., Kawano, F., Niimi, M., Tobinai, K., Hotta, T., & Shimoyama, M. (2001). A new G-CSF-supported combination chemotherapy, LSG15, for adult T-cell leukaemia-lymphoma: Japan Clinical Oncology Group Study 9303. British journal of haematology, 113(2), 375–382. [CrossRef]
- Tsukasaki, K., Tobinai, K., Shimoyama, M., Kozuru, M., Uike, N., Yamada, Y., Tomonaga, M., Araki, K., Kasai, M., Takatsuki, K., Tara, M., Mikuni, C., Hotta, T., & Lymphoma Study Group of the Japan Clinical Oncology Group (2003). Deoxycoformycin-containing combination chemotherapy for adult T-cell leukemia-lymphoma: Japan Clinical Oncology Group Study (JCOG9109). International journal of hematology, 77(2), 164–170. [CrossRef]
- Ishida, T., Jo, T., Takemoto, S., Suzushima, H., Uozumi, K., Yamamoto, K., Uike, N., Saburi, Y., Nosaka, K., Utsunomiya, A., Tobinai, K., Fujiwara, H., Ishitsuka, K., Yoshida, S., Taira, N., Moriuchi, Y., Imada, K., Miyamoto, T., Akinaga, S., Tomonaga, M., … Ueda, R. (2015). Dose-intensified chemotherapy alone or in combination with mogamulizumab in newly diagnosed aggressive adult T-cell leukaemia-lymphoma: a randomized phase II study. British journal of haematology, 169(5), 672–682. [CrossRef]
- Horwitz, S., O'Connor, O. A., Pro, B., Illidge, T., Fanale, M., Advani, R., Bartlett, N. L., Christensen, J. H., Morschhauser, F., Domingo-Domenech, E., Rossi, G., Kim, W. S., Feldman, T., Lennard, A., Belada, D., Illés, Á., Tobinai, K., Tsukasaki, K., Yeh, S. P., Shustov, A., … ECHELON-2 Study Group (2019). Brentuximab vedotin with chemotherapy for CD30-positive peripheral T-cell lymphoma (ECHELON-2): a global, double-blind, randomised, phase 3 trial. Lancet (London, England), 393(10168), 229–240. [CrossRef]
- Horwitz, S., O'Connor, O. A., Pro, B., Trümper, L., Iyer, S., Advani, R., Bartlett, N. L., Christensen, J. H., Morschhauser, F., Domingo-Domenech, E., Rossi, G., Kim, W. S., Feldman, T., Menne, T., Belada, D., Illés, Á., Tobinai, K., Tsukasaki, K., Yeh, S. P., Shustov, A., … Illidge, T. (2022). The ECHELON-2 Trial: 5-year results of a randomized, phase III study of brentuximab vedotin with chemotherapy for CD30-positive peripheral T-cell lymphoma. Annals of oncology : official journal of the European Society for Medical Oncology, 33(3), 288–298. [CrossRef]
- Ishida, T., Fujiwara, H., Nosaka, K., Taira, N., Abe, Y., Imaizumi, Y., Moriuchi, Y., Jo, T., Ishizawa, K., Tobinai, K., Tsukasaki, K., Ito, S., Yoshimitsu, M., Otsuka, M., Ogura, M., Midorikawa, S., Ruiz, W., & Ohtsu, T. (2016). Multicenter Phase II Study of Lenalidomide in Relapsed or Recurrent Adult T-Cell Leukemia/Lymphoma: ATLL-002. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 34(34), 4086–4093. [CrossRef]
- Bazarbachi, A., Plumelle, Y., Carlos Ramos, J., Tortevoye, P., Otrock, Z., Taylor, G., Gessain, A., Harrington, W., Panelatti, G., & Hermine, O. (2010). Meta-analysis on the use of zidovudine and interferon-alfa in adult T-cell leukemia/lymphoma showing improved survival in the leukemic subtypes. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 28(27), 4177–4183. [CrossRef]
- Macchi, B., Balestrieri, E., Frezza, C., Grelli, S., Valletta, E., Marçais, A., Marino-Merlo, F., Turpin, J., Bangham, C. R., Hermine, O., Mastino, A., & Bazarbachi, A. (2017). Quantification of HTLV-1 reverse transcriptase activity in ATL patients treated with zidovudine and interferon-α. Blood advances, 1(12), 748–752. [CrossRef]
- Hleihel, R., Akkouche, A., Skayneh, H., Hermine, O., Bazarbachi, A., & El Hajj, H. (2021). Adult T-Cell Leukemia: a Comprehensive Overview on Current and Promising Treatment Modalities. Current oncology reports, 23(12), 141. [CrossRef]
- Bazarbachi, A., El-Sabban, M. E., Nasr, R., Quignon, F., Awaraji, C., Kersual, J., Dianoux, L., Zermati, Y., Haidar, J. H., Hermine, O., & de Thé, H. (1999). Arsenic trioxide and interferon-alpha synergize to induce cell cycle arrest and apoptosis in human T-cell lymphotropic virus type I-transformed cells. Blood, 93(1), 278–283.
- Mahieux, R., Pise-Masison, C., Gessain, A., Brady, J. N., Olivier, R., Perret, E., Misteli, T., & Nicot, C. (2001). Arsenic trioxide induces apoptosis in human T-cell leukemia virus type 1- and type 2-infected cells by a caspase-3-dependent mechanism involving Bcl-2 cleavage. Blood, 98(13), 3762–3769. [CrossRef]
- El Hajj, H., El-Sabban, M., Hasegawa, H., Zaatari, G., Ablain, J., Saab, S. T., Janin, A., Mahfouz, R., Nasr, R., Kfoury, Y., Nicot, C., Hermine, O., Hall, W., de Thé, H., & Bazarbachi, A. (2010). Therapy-induced selective loss of leukemia-initiating activity in murine adult T cell leukemia. The Journal of experimental medicine, 207(13), 2785–2792. [CrossRef]
- El-Sabban, M. E., Nasr, R., Dbaibo, G., Hermine, O., Abboushi, N., Quignon, F., Ameisen, J. C., Bex, F., de Thé, H., & Bazarbachi, A. (2000). Arsenic-interferon-alpha-triggered apoptosis in HTLV-I transformed cells is associated with tax down-regulation and reversal of NF-kappa B activation. Blood, 96(8), 2849–2855.
- Kchour, G., Tarhini, M., Kooshyar, M. M., El Hajj, H., Wattel, E., Mahmoudi, M., Hatoum, H., Rahimi, H., Maleki, M., Rafatpanah, H., Rezaee, S. A., Yazdi, M. T., Shirdel, A., de Thé, H., Hermine, O., Farid, R., & Bazarbachi, A. (2009). Phase 2 study of the efficacy and safety of the combination of arsenic trioxide, interferon alpha, and zidovudine in newly diagnosed chronic adult T-cell leukemia/lymphoma (ATL). Blood, 113(26), 6528–6532. [CrossRef]
- Marçais, A., Cook, L., Witkover, A., Asnafi, V., Avettand-Fenoel, V., Delarue, R., Cheminant, M., Sibon, D., Frenzel, L., de Thé, H., Bangham, C. R. M., Bazarbachi, A., Hermine, O., & Suarez, F. (2020). Arsenic trioxide (As2O3) as a maintenance therapy for adult T cell leukemia/lymphoma. Retrovirology, 17(1), 5. [CrossRef]
- Kchour, G., Rezaee, R., Farid, R., Ghantous, A., Rafatpanah, H., Tarhini, M., Kooshyar, M. M., El Hajj, H., Berry, F., Mortada, M., Nasser, R., Shirdel, A., Dassouki, Z., Ezzedine, M., Rahimi, H., Ghavamzadeh, A., de Thé, H., Hermine, O., Mahmoudi, M., & Bazarbachi, A. (2013). The combination of arsenic, interferon-alpha, and zidovudine restores an "immunocompetent-like" cytokine expression profile in patients with adult T-cell leukemia lymphoma. Retrovirology, 10, 91. [CrossRef]
- Moura, I. C., Lepelletier, Y., Arnulf, B., England, P., Baude, C., Beaumont, C., Bazarbachi, A., Benhamou, M., Monteiro, R. C., & Hermine, O. (2004). A neutralizing monoclonal antibody (mAb A24) directed against the transferrin receptor induces apoptosis of tumor T lymphocytes from ATL patients. Blood, 103(5), 1838–1845. [CrossRef]
- Sharma, K., Janik, J. E., O'Mahony, D., Stewart, D., Pittaluga, S., Stetler-Stevenson, M., Jaffe, E. S., Raffeld, M., Fleisher, T. A., Lee, C. C., Steinberg, S. M., Waldmann, T. A., & Morris, J. C. (2017). Phase II Study of Alemtuzumab (CAMPATH-1) in Patients with HTLV-1-Associated Adult T-cell Leukemia/lymphoma. Clinical cancer research : an official journal of the American Association for Cancer Research, 23(1), 35–42. [CrossRef]
- Zhang, Z., Zhang, M., Ravetch, J. V., Goldman, C., & Waldmann, T. A. (2003). Effective therapy for a murine model of adult T-cell leukemia with the humanized anti-CD2 monoclonal antibody, MEDI-507. Blood, 102(1), 284–288. [CrossRef]
- Waldmann, T. A., White, J. D., Goldman, C. K., Top, L., Grant, A., Bamford, R., Roessler, E., Horak, I. D., Zaknoen, S., & Kasten-Sportes, C. (1993). The interleukin-2 receptor: a target for monoclonal antibody treatment of human T-cell lymphotrophic virus I-induced adult T-cell leukemia. Blood, 82(6), 1701–1712.
- Baba, Y., Sakai, H., Kabasawa, N., & Harada, H. (2023). Successful Treatment of an Aggressive Adult T-cell Leukemia/Lymphoma with Strong CD30 Expression Using Brentuximab Vedotin as Combination and Maintenance Therapy. Internal medicine (Tokyo, Japan), 62(4), 613–616. [CrossRef]
- Iqbal, M., Reljic, T., Klocksieben, F., Sher, T., Ayala, E., Murthy, H., Bazarbachi, A., Kumar, A., & Kharfan-Dabaja, M. A. (2019). Efficacy of Allogeneic Hematopoietic Cell Transplantation in Human T Cell Lymphotropic Virus Type 1-Associated Adult T Cell Leukemia/Lymphoma: Results of a Systematic Review/Meta-Analysis. Biology of blood and marrow transplantation : journal of the American Society for Blood and Marrow Transplantation, 25(8), 1695–1700. [CrossRef]
- Fukushima, T., Miyazaki, Y., Honda, S., Kawano, F., Moriuchi, Y., Masuda, M., Tanosaki, R., Utsunomiya, A., Uike, N., Yoshida, S., Okamura, J., & Tomonaga, M. (2005). Allogeneic hematopoietic stem cell transplantation provides sustained long-term survival for patients with adult T-cell leukemia/lymphoma. Leukemia, 19(5), 829–834. [CrossRef]
- Hishizawa, M., Kanda, J., Utsunomiya, A., Taniguchi, S., Eto, T., Moriuchi, Y., Tanosaki, R., Kawano, F., Miyazaki, Y., Masuda, M., Nagafuji, K., Hara, M., Takanashi, M., Kai, S., Atsuta, Y., Suzuki, R., Kawase, T., Matsuo, K., Nagamura-Inoue, T., Kato, S., … Uchiyama, T. (2010). Transplantation of allogeneic hematopoietic stem cells for adult T-cell leukemia: a nationwide retrospective study. Blood, 116(8), 1369–1376. [CrossRef]
- Nakamura, T., Oku, E., Nomura, K., Morishige, S., Takata, Y., Seki, R., Imamura, R., Osaki, K., Hashiguchi, M., Yakushiji, K., Mouri, F., Mizuno, S., Yoshimoto, K., Ohshima, K., Nagafuji, K., & Okamura, T. (2012). Unrelated cord blood transplantation for patients with adult T-cell leukemia/lymphoma: experience at a single institute. International journal of hematology, 96(5), 657–663. [CrossRef]
- Kato, K., Choi, I., Wake, A., Uike, N., Taniguchi, S., Moriuchi, Y., Miyazaki, Y., Nakamae, H., Oku, E., Murata, M., Eto, T., Akashi, K., Sakamaki, H., Kato, K., Suzuki, R., Yamanaka, T., & Utsunomiya, A. (2014). Treatment of patients with adult T cell leukemia/lymphoma with cord blood transplantation: a Japanese nationwide retrospective survey. Biology of blood and marrow transplantation : journal of the American Society for Blood and Marrow Transplantation, 20(12), 1968–1974. [CrossRef]
- Tanase, A. D., Colita, A., Craciun, O. G., Lipan, L., Varady, Z., Stefan, L., Ranete, A., Pasca, S., Bumbea, H., Andreescu, M., Popov, V., Bardas, A., Coriu, D., Lupu, A. R., Tomuleasa, C., Colita, A., & Hermine, O. (2020). Allogeneic Stem Cell Transplantation for Adult T-Cell Leukemia/Lymphoma-Romanian Experience. Journal of clinical medicine, 9(8), 2417. [CrossRef]
- Nakata, K., Fuji, S., Koike, M., Tada, Y., Masaie, H., Yoshida, H., Watanabe, E., Kobayashi, S., Tojo, A., Uchimaru, K., & Ishikawa, J. (2020). Successful treatment strategy in corporating mogamulizumab and cord blood transplantation in aggressive adult T-cell leukemia-lymphoma: a case report. Blood cell therapy, 3(1), 6–10. [CrossRef]
- Takeda, A., Sakoda, T., Yawata, N., Kato, K., Hasegawa, E., Shima, T., Hikita, S., Yoshitomi, K., Takenaka, K., Oda, Y., Akashi, K., & Sonoda, K. H. (2022). Panuveitis induced by donor-derived CD8+ T cells after allogeneic hematopoietic stem cell transplantation for adult T-cell leukemia. American journal of ophthalmology case reports, 27, 101673. [CrossRef]
- Margueron, R., & Reinberg, D. (2011). The Polycomb complex PRC2 and its mark in life. Nature, 469(7330), 343–349. [CrossRef]
- Fujikawa, D., Nakagawa, S., Hori, M., Kurokawa, N., Soejima, A., Nakano, K., Yamochi, T., Nakashima, M., Kobayashi, S., Tanaka, Y., Iwanaga, M., Utsunomiya, A., Uchimaru, K., Yamagishi, M., & Watanabe, T. (2016). Polycomb-dependent epigenetic landscape in adult T-cell leukemia. Blood, 127(14), 1790–1802. [CrossRef]
- Sasaki, D., Imaizumi, Y., Hasegawa, H., Osaka, A., Tsukasaki, K., Choi, Y. L., Mano, H., Marquez, V. E., Hayashi, T., Yanagihara, K., Moriwaki, Y., Miyazaki, Y., Kamihira, S., & Yamada, Y. (2011). Overexpression of Enhancer of zeste homolog 2 with trimethylation of lysine 27 on histone H3 in adult T-cell leukemia/lymphoma as a target for epigenetic therapy. Haematologica, 96(5), 712–719. [CrossRef]
- Izutsu, K., Makita, S., Nosaka, K., Yoshimitsu, M., Utsunomiya, A., Kusumoto, S., Morishima, S., Tsukasaki, K., Kawamata, T., Ono, T., Rai, S., Katsuya, H., Ishikawa, J., Yamada, H., Kato, K., Tachibana, M., Kakurai, Y., Adachi, N., Tobinai, K., Yonekura, K., … Ishitsuka, K. (2023). An open-label, single-arm phase 2 trial of valemetostat for relapsed or refractory adult T-cell leukemia/lymphoma. Blood, 141(10), 1159–1168. [CrossRef]
- El Hajj, H., Hermine, O., & Bazarbachi, A. (2024). Therapeutic advances for the management of adult T cell leukemia: Where do we stand?. Leukemia research, 147, 107598. [CrossRef]
- Nakamura, H., Hayami, M., Ohta, Y., Ishikawa, K., Tsujimoto, H., Kiyokawa, T., Yoshida, M., Sasagawa, A., & Honjo, S. (1987). Protection of cynomolgus monkeys against infection by human T-cell leukemia virus type-I by immunization with viral env gene products produced in Escherichia coli. International journal of cancer, 40(3), 403–407. [CrossRef]
- Hakoda, E., Machida, H., Tanaka, Y., Morishita, N., Sawada, T., Shida, H., Hoshino, H., & Miyoshi, I. (1995). Vaccination of rabbits with recombinant vaccinia virus carrying the envelope gene of human T-cell lymphotropic virus type I. International journal of cancer, 60(4), 567–570. [CrossRef]
- Franchini, G., Tartaglia, J., Markham, P., Benson, J., Fullen, J., Wills, M., Arp, J., Dekaban, G., Paoletti, E., & Gallo, R. C. (1995). Highly attenuated HTLV type Ienv poxvirus vaccines induce protection against a cell-associated HTLV type I challenge in rabbits. AIDS research and human retroviruses, 11(2), 307–313. [CrossRef]
- Ibuki, K., Funahashi, S. I., Yamamoto, H., Nakamura, M., Igarashi, T., Miura, T., Ido, E., Hayami, M., & Shida, H. (1997). Long-term persistence of protective immunity in cynomolgus monkeys immunized with a recombinant vaccinia virus expressing the human T cell leukaemia virus type I envelope gene. The Journal of general virology, 78 ( Pt 1), 147–152. [CrossRef]
- Armand, M. A., Grange, M. P., Paulin, D., & Desgranges, C. (2000). Targeted expression of HTLV-I envelope proteins in muscle by DNA immunization of mice. Vaccine, 18(21), 2212–2222. [CrossRef]
- Kazanji, M., Heraud, J. M., Merien, F., Pique, C., de Thé, G., Gessain, A., & Jacobson, S. (2006). Chimeric peptide vaccine composed of B- and T-cell epitopes of human T-cell leukemia virus type 1 induces humoral and cellular immune responses and reduces the proviral load in immunized squirrel monkeys (Saimiri sciureus). The Journal of general virology, 87(Pt 5), 1331–1337. [CrossRef]
- Suehiro, Y., Hasegawa, A., Iino, T., Sasada, A., Watanabe, N., Matsuoka, M., Takamori, A., Tanosaki, R., Utsunomiya, A., Choi, I., Fukuda, T., Miura, O., Takaishi, S., Teshima, T., Akashi, K., Kannagi, M., Uike, N., & Okamura, J. (2015). Clinical outcomes of a novel therapeutic vaccine with Tax peptide-pulsed dendritic cells for adult T cell leukaemia/lymphoma in a pilot study. British journal of haematology, 169(3), 356–367. [CrossRef]
- Sugata, K., Yasunaga, J., Mitobe, Y., Miura, M., Miyazato, P., Kohara, M., & Matsuoka, M. (2015). Protective effect of cytotoxic T lymphocytes targeting HTLV-1 bZIP factor. Blood, 126(9), 1095–1105. [CrossRef]
- Daian E Silva, D. S. O., Cox, L. J., Rocha, A. S., Lopes-Ribeiro, Á., Souza, J. P. C., Franco, G. M., Prado, J. L. C., Pereira-Santos, T. A., Martins, M. L., Coelho-Dos-Reis, J. G. A., Gomes-de-Pinho, T. M., Da Fonseca, F. G., & Barbosa-Stancioli, E. F. (2023). Preclinical assessment of an anti-HTLV-1 heterologous DNA/MVA vaccine protocol expressing a multiepitope HBZ protein. Virology journal, 20(1), 304. [CrossRef]
- Tu, J. J., King, E., Maksimova, V., Smith, S., Macias, R., Cheng, X., Vegesna, T., Yu, L., Ratner, L., Green, P. L., Niewiesk, S., Richner, J. M., & Panfil, A. R. (2024). An HTLV-1 envelope mRNA vaccine is immunogenic and protective in New Zealand rabbits. Journal of virology, 98(2), e0162323. [CrossRef]
- Ishitsuka K. (2021). Diagnosis and management of adult T-cell leukemia/lymphoma. Seminars in hematology, 58(2), 114–122. [CrossRef]
Figure 1.
Primary modes of viral spread.
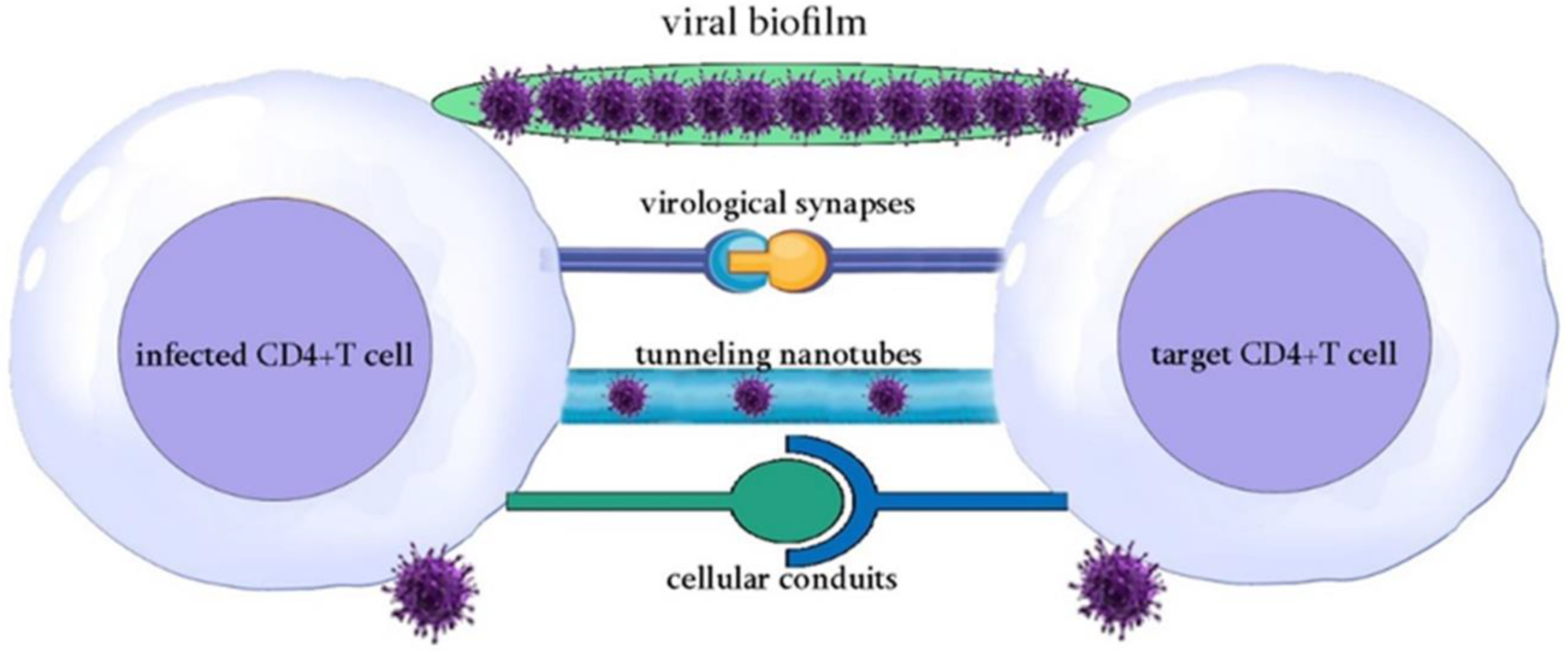
Figure 2.
HTLV-1 replication cycle in Host Cell.
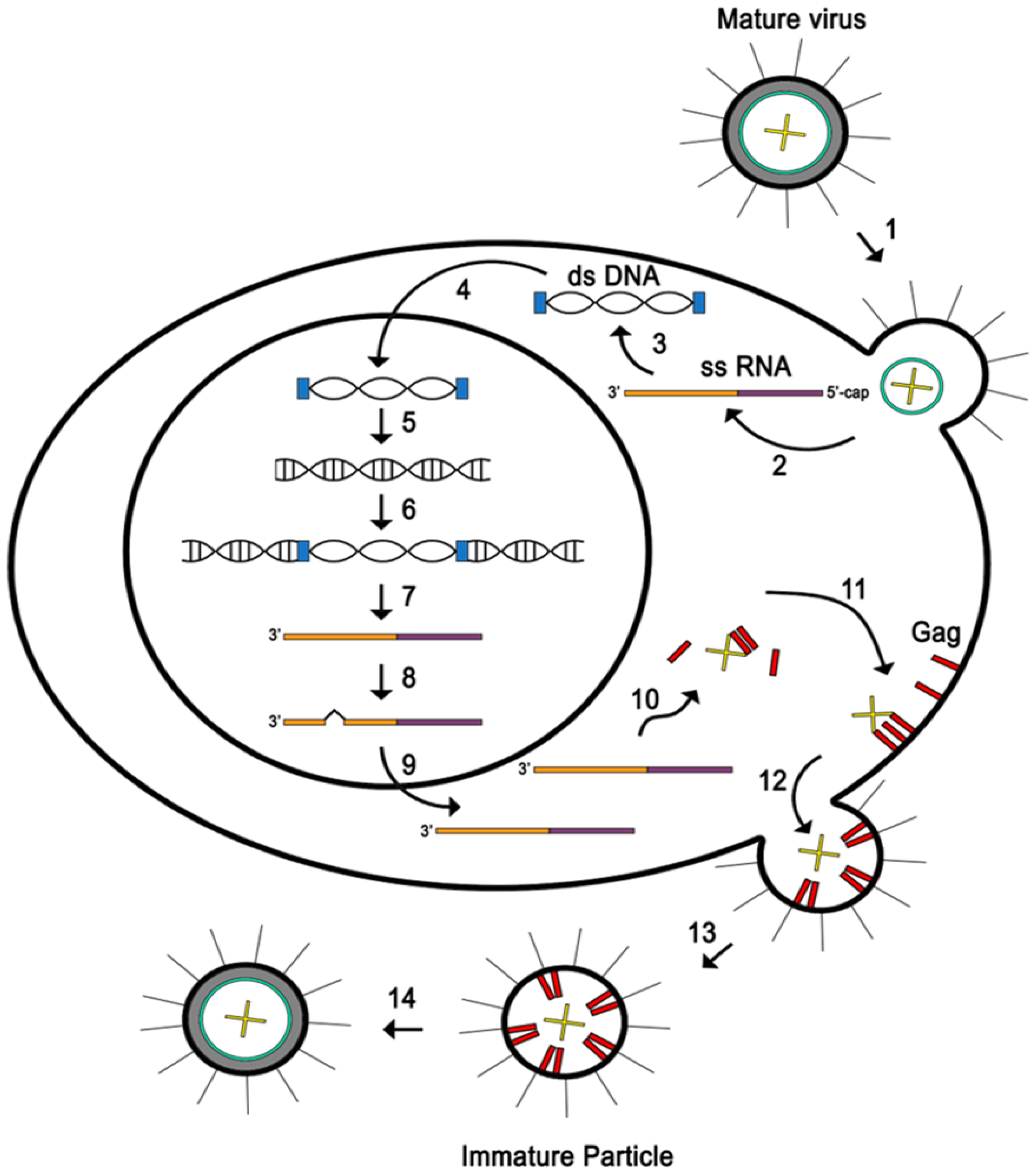

Table 1.
Combination chemotherapy used to treat adult T-cell leukaemia-lymphoma in the clinical setting.
Table 1.
Combination chemotherapy used to treat adult T-cell leukaemia-lymphoma in the clinical setting.
| Study | Treatment regimen | Cinical endpoints | Adverse effects | Disease | References |
|---|---|---|---|---|---|
| A multiinstitutional, cooperative study (1996) | CHOP (cyclophosphamide, doxorubicin, vincristine, and prednisone) followed by etoposide, vindesine, ranimustine, and mitoxantrone supported by the granulocyte colony-stimulating factor (G-CSF) | Complete remission rate of 35.8%, partial remission rate of 38.3%, median survival of 8.5 months, predicted 3-year survival of 13.5% | Adult T-cell leukemia/lymphoma | [28] | |
| A multicenter phase II study (2001) | VCAP (vincristine, cyclophosphamide, doxorubicin and prednisone), AMP (doxorubicin, ranimustine and prednisone) and VECP (vindesine, etoposide, carboplatin and prednisone), G-CSF was administered during the intervals between chemotherapy | Complete response rate of 35.5%, partial response rate of 45.2%, median survival time of 13 months, estimated 2-year overall survival of 31.3% | Grade 4 hematological toxicity of neutropenia in 65.3% of patients, thrombocytopenia in 52.6% of patients. | Adult T-cell leukemia-lymphoma | [29] |
| A multicenter phase II study (2003) |
Vincristine, doxorubicin, etoposide, prednisolone, and Deoxycoformycin (an inhibitor of adenosine deaminase) | Complete response of 28%, partial response of 24%, median survival time of 7.4 months, estimated 2-year survival rate of 15.5% | Grade 4 neutropenia in 67%, grade 3 or greater infection in 22%, treatment-related death in 7% (4 patients), septicemia in 2 patients, cytomegalovirus pneumonia in 2 patients. This combination chemotherapy is not a promising regimen against aggressive ATL. | Adult T-cell leukemia-lymphoma | [30] |
| A multicentre, randomized, phase II study (2015) | VCAP-AMP-VECP | Complete response rate of 33%, overall response rate of 77% | Adult T-cell leukemia-lymphoma |
[31] | |
| VCAP-AMP-VECP plus mogamulizumab (antibody targeting CC chemokine receptor type 4 ) | Complete response rate of 52%, overall response rate of 86% | Grade ≥3 treatment-emergent adverse events including anemia, thrombocytopenia, lymphopenia, leukopenia, and decreased appetite, additional adverse events including skin disorders, cytomegalovirus infection, pyrexia, hyperglycemia, and interstitial lung disease | |||
| A global, double-blind, randomised, phase 3 trial (2019, 2022) |
Brentuximab vedotin (anti-CD30 antibody), cyclophosphamide, doxorubicin, and prednisone | Median progression-free survival of 48,2 months, 5-year progression-free survival rate of 51.4%, 5-year overall survival rate of 70.1% | Adverse events, including incidence and severity of febrile neutropenia in 18% of patients and peripheral neuropathy in 52% of patients. Fatal adverse events occurred in 3% of patients | CD30-positive peripheral T-cell lymphomas | [32,33] |
| CHOP | Median progression-free survival of 20,8 months, 5-year progression-free survival rates of 43%, 5-year overall survival (OS) rate of 61% | Adverse events, including incidence and severity of febrile neutropenia in 15% of patients in and peripheral neuropathy in 55% of the patients. Fatal adverse events occurred in 4% of the patients. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



