
Submitted:
28 February 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Understanding how organisms respond to thermal stressors over short- and long-term timescales is essential for assessing and predicting their resilience and vulnerability to environmental changes and climate fluctuations. Here, using a biparental-care burying beetle (Nicrophorus vespilloides), we tested whether and how parental investment in carcass preparation and thermal condition interact to influence subsequent parental care behaviour and reproductive success. We employed a 3 × 2 factorial experiment, manipulating the levels of parental investment in carcass preparation (Reduced, Control, and Elevated) and ambient temperatures (benign: 20°C and harsh: 23°C) in breeding pairs. We found that (1) irrespective of ambient temperature, males in the Reduced group decreased their pre-hatching care. (2) Across all investment groups, both sexes under the higher temperature reduced post-hatching care. (3) Carcass-preparation investment and thermal condition interactively influenced reproductive success. Overall, the harsh temperature decreased reproductive success. Furthermore, beetle pairs experienced reduced carcass-preparation investment producing fewer eggs and lighter broods, while those experienced elevated carcass-preparation investment producing smaller and lighter broods. Our findings provide new insights into how thermal stress affects parental investment strategies, and enhance our understanding of the phenotypic plasticity in reproductive strategies that animals employ to cope with climate change.
Keywords:
burying beetle
; climate change
; life-history trade-offs
; parental investment
1. Introduction
Global warming, driven by climate change, is a major environmental and ecological challenge with profound impacts on biodiversity [1]. It severely affects individual survival and animal populations through both lethal and non-lethal mechanisms [2], such as thermal changes impairing animal reproduction [3,4,5,6]. Numerous experimental studies across animal taxa have investigated the impact of ambient temperature on life-history traits and reproductive performance and behaviour, such as sperm performance [7,8,9], fecundity [8,10], hatching success [11,12], parental care [13,14,15], and offspring performance [14,15]. These thermal effects may not only act directly on these traits but also indirectly by altering external factors that influence them. For example, rising temperatures and increased frequency of extreme events can shift the phenology of plants and insects, which subsequently affects the timing of birds’ migration [16] and reproduction [17]. Alternatively, temperature changes can directly affect reproductive behaviours, such as the regulation of egg-laying time in great tits (Parus major) [18], and offspring fitness in tree swallows (Tachycineta bicolor) [19]. Importantly, the impacts of thermal conditions are often not limited to immediate effects but extend to long-term consequences as well [20]. For example, in the Chinese lacertid lizard (Takydromus septentrionalis), offspring of parents exposed to elevated thermal conditions exhibited higher survival rates under simulated future climate scenarios, but not under current climatic conditions [21]. Another recent finding suggests that female burying beetles (Nicrophorus vespilloides) exposed to heatwave during the pupal stage are more likely to provide enhanced direct care for their offspring [22]. Given these insights, it is crucial to consider other relevant factors, such as animal groups and resource availability, when investigating the thermal effects on individual fitness. Furthermore, understanding how organisms respond to thermal stressor across short- and long-term timescales is essential for predicting their resilience and vulnerability to climate change.
Despite extensive research on the effects of thermal stress on the reproductive success [23], the interactions between temperature and other ecological factors in shaping life-history traits and behaviours remain less studied. Life history theory predicts trade-offs (i.e., negative relationship) among different life-history traits [24], such as offspring number versus offspring size [25], or current reproductive investment versus future survival and reproductive opportunities [26,27]. Parental investment involves the allocation of resources toward offspring at the current reproduction at the expense of future reproductive potential [28]. These strategies are shaped by intrinsic factors (e.g., prior reproductive experience, energy reserves) and extrinsic environmental conditions (e.g., resource availability, thermal conditions). Specifically, parental investment theory suggests that individuals adjust their current reproductive investment based on their past parental investment [29]. Resource availability influences energy stores for parents, which in turn regulate trade-offs between different life-history traits and behaviours [30,31]. For example, when resources are abundant, some studies report weak or even positive correlations between traits, suggesting that trade-offs may be context-dependent [32,33]. Furthermore, ambient temperature modulates metabolic rates in both ectothermic and endothermic animals, with cascading effects on physiological processes and behaviours [34]. However, knowledge gaps remain regarding the impact of thermal stress on reproduction and the adjustment of individual parental effort within and between reproductive events. Addressing these gaps will enhance our understanding of how thermal challenges modulate parental investment strategies and contribute to phenotypic plasticity in reproductive behaviours as adaptive responses to climate-driven environmental pressures. In this study, we investigate how parental investment (i.e., investment in processing resources) influences subsequent parental care behaviour and investment (both pre- and post-hatching) under varying thermal conditions in the burying beetles (N. vespilloides). Given its well-documented reproductive traits and extensively studied parental care behaviour, the burying beetle serves as an ideal model to explore the reproductive adaptation of animals in response to environmental changes.
Burying beetles (Nicrophorus spp.) are scavenger insects that exploit small vertebrate carcasses as both food and breeding resources. These beetles exhibit complex biparental care, with both males and females cooperatively engaging in carcass preparation and maintenance, brood defense, and food provisioning for offspring [35]. Parental care occurs in two distinct phases: pre-hatching and post-hatching, separated by egg hatching. Pre-hatching care primarily involves beetles processing carcasses by removing any fur or feathers, rolling it into a ball, and smearing it with oral and anal secretions [36,37,38,39]. Post-hatching care consists of feeding developing larvae with regurgitated and predigested food (i.e., direct care), as well as additional behaviours associated with carcass maintenance and defense (i.e., indirect care). Vertebrate carcasses are ephemeral and limited resources, making burying beetles opportunistic breeders that can reproduce multiple times throughout their lifetime [35]. Both male and female beetles differentially adjust their reproductive strategies in response to various factors, such as age [40], inbreeding [41], brood size [42], nutritional status [43,44], carcass size [45], and breeding patterns (i.e., cobreeding or communal breeding, pair breeding, and male and female uniparental breeding) [46,47,48,49,50,51], and temperature changes [6,13,14]. Some studies have demonstrated the impact of thermal variation on reproductive success and parental care behaviour in burying beetles [6,13,14,52]. For instance, in the burying beetle (Nicrophorus orbicollis), thermal stress significantly reduces egg number, fertilization success, dispersed larval count, and mean larval mass, while parental care behaviors in both sexes remain unaffected [13]. However, whether and how thermal variation modulates parental investment and its trade-offs within and across breeding events remains poorly understood.
To address this gap, we employed a 3 × 2 factorial experimental design to explore whether and how breeding pairs of beetles adjust their parental care behaviour and investment in response to variation in carcass-preparation effort within one breeding event, as well as how ambient temperature influences these parental adjustments. We manipulated three levels of parental investment in carcass preparation: Reduced, Control, and Elevated treatments. In the reduced-investment group (Reduced group), beetle pairs were provided with prepared carcasses of mice, simulating low investment in carcass preparation for beetles. As the Control group, beetle pairs were provided with newly thawed frozen carcasses of mice. To simulate a heightened-investment scenario, we also created the elevated-investment group (Elevated group), where beetle pairs have prepared carcasses before (i.e., with prior carcass-preparation experience), and subsequently were given freshly thawed carcasses for breeding. Furthermore, beetles were exposed to two thermal conditions (20 °C and 23 °C) throughout the whole period of breeding events, including the carcass preparation process. The 20 °C condition was considered benign, as the beetles used in this study were six-generation descendants reared under this temperature. In contrast, the 23 °C condition was deemed harsh, as it imposes reproductive costs on the beetles [6,13]. A pilot experiment demonstrated that 25 °C led to severe reproductive failure (0/120 broods), suggesting the 23 °C condition was stressful but not immediately detrimental to reproduction. To assess the effects of carcass-preparation investment and breeding temperature on individual care strategies and reproductive success, we measured pre- and post-hatching care behaviours, metrics of reproductive success (e.g., clutch size, brood size, brood mass, and offspring performance (i.e., mean larval mass)). We also calculated the relative change in parental body mass to assess the parental performance after breeding.
Carcass preparation contains a series of energetically costly behaviours that may limit the parent’s ability to invest in their offspring [53,54], potentially reducing reproductive success. Additionally, the rate of carcass decomposition varies across ambient thermal conditions [55], which may lead to burying beetle parents to adjust their time and energy during carcass preparation, thereby affecting resource allocations for subsequent investments. For instance, under higher temperatures, parents may expend more effort on carcass preparation, limiting resources available for offspring care and reducing reproductive success. Thus, we predicted that beetles in the Reduced group would exhibit increased parental care and higher reproductive success, with this effect being more pronounced under benign thermal conditions. Conversely, we expected beetles in the Elevated group to show decreased parental care and lower reproductive success, whereas harsh thermal conditions may further exacerbate these negative effects on parental care and reproduction. Our study will illuminate how reproductive allocation within breeding events is influenced by thermal variations. By doing so, it will enhance our understanding of the phenotypic plasticity in parental investment strategies that animals, and insects in particular, potentially employ to adapt to the challenges posed by climate change.
2. Materials and Methods
2.1. Beetles Husbandry
All adult beetles in this study were sixth-generation descendants from an outbred laboratory population at the University of Groningen, originally sourced from a wild population in De Vosbergen, The Netherlands (53 ° 08 ' N, 06 ° 35 ' E) in the summer of 2022. Up to six same-sex adults from the same brood were housed together in clear rearing boxes (10 cm L × 6 cm W × 8 cm H) with moist peat and fed two beheaded mealworms (Tenebrio molitor) per beetle twice weekly. The husbandry room was set at 20°C with a 16 h: 8 h light cycle.
2.2. Experimental Protocol
We established three carcass-preparation investment groups (Reduced, Control, and Elevated) and conducted the experiment under two distinct ambient thermal conditions (20 °C and 23 °C) independently (Figure 1). In the Reduced group (20 °C: n = 30; 23 °C: n = 30), beetle pairs that did not undergo carcass preparation were provided with prepared carcasses of mice as breeding resources. In the Control group (20 °C: n = 63; 23 °C: n = 56), beetle pairs that did not undergo carcass preparation were provided with freshly thawed dead mice as breeding resources. In the Elevated group (20 °C: n = 28; 23 °C: n = 25), beetle pairs that performed carcass preparation once were given freshly thawed dead mice as breeding resources.
Non-sibling, virgin, sexually mature (ca. 2 weeks post-eclosion) males (mean ± SE = 5.11 ± 0.02 mm) and size-matched females (mean ± SE = 5.05 ± 0.02 mm) were paired one day before initiating breeding. This pairing method minimized the potential effects of inbreeding and individual reproductive history on reproductive success, and ensured that females laid fertilized eggs [56,57]. Prior to pairing, we measured each beetle's body size (i.e., middle width of the pronotum) using an electric caliper (accuracy: 0.01 mm). Then each pair of beetles was transferred into a breeding container (23 cm L × 19 cm W × 12.5 cm H) filled with 3–5 cm of moist peat and supplied a mouse carcass (prepared carcasses of mice for the Reduced group; thawed, unprepared carcasses for the Control and Elevated groups: mean ± SE = 23.71 ± 0.18 g) to initiate breeding. To obtain prepared carcasses for the Reduced groups, we conducted preliminary breeding trials before the main experiment. In these trails, beetle pairs were each given a carcass for preparation at either 20 °C or 23 °C. After approximately 60 hours, prepared carcasses (i.e., approximately 90 % of fur was removed, and the body was shaped into a ball) were collected and stored at 4 °C for later use [57], whereas the beetle pairs that have prepared carcasses were transferred into new breeding containers for use in the Elevated groups. Specifically, during the carcass preparation period (i.e., pre-hatching period, before egg hatching), beetle pairs in the Reduced group only need to drag the prepared carcass into the soil, bury it and add some oral and anal secretions to the carcass surface, but are released from hard work of removing the furs and rolling the carcass into a ball shape. Prior to initiating breeding events, we also weighed the body mass of beetles using an analytical balance (accuracy: 0.0001 g). Additionally, all individuals were marked with small holes in the elytra (i.e., left one for males and right one for females) using a 00-insect pin to facilitate individual recognition during subsequent daily inspections.
After beetle pairs were given carcasses for breeding, we conducted visual inspections of the breeding containers three times daily (between 07:00–08:30 am, 1:30–3:00 pm, 8:00–10:30 pm, at 5-hour intervals) until breeding success which is defined as at least one developing larva leaving from the carcass (i.e., larval dispersal) [51]. At each inspection, we slightly removed the surface soil of the carcass to observe whether and which parent(s) was (were) present on or within the carcass. Parental presence was defined as parental care no matter what actual behaviours the parents do (e.g., removing fur, rolling the carcass, feeding larvae, alert, cleaning the antennae and tarsus, and chewing carrion), while parental absence was defined as no parental care [48,51,57]. Before egg hatching, we also counted the number of visible eggs at the bottom of the breeding containers during daily inspections. The highest egg count observed during these checks was recorded as the clutch size [14,58,59]. At the larval dispersal stage, we enumerated all dispersed larvae to determine brood size and weighed them using an analytical balance (accuracy: 0.0001 g) to quantify brood mass, which provides estimates of reproductive success. Furthermore, we calculated mean larval mass, an estimate of offspring fitness, by dividing the brood mass by the brood size. Additionally, we measured the body mass of surviving parents at the larval dispersal. The change in parental body mass during a breeding event was used as an indicator of residual reproductive value and calculated using the following formula: Body mass change = (Mass at larval dispersal − Mass before breeding) / Mass before breeding [48,51,57]. Breeding events without dispersed larvae or with dead parental individuals were excluded from our dataset.
Parental care across two reproductive stages—pre-hatching and post-hatching—was quantified separately using the following equations: (1) Pre-hatching care = (number of observations of the focal parent present on or within the carcass prior to the emergence of the first larva) / (total number of daily inspections conducted before larval emergence); (2) Post-hatching care = (number of observations of the focal parent present on or within the carcass after the emergence of the first larva) / (total number of daily inspections conducted after larval emergence). These calculations provide a comparative measure of parental investment during distinct developmental stages [48,51,57]. Meanwhile, parental care during the whole breeding phase was also calculated using the equation: parental care = (number of observations of the focal parent present on or within the carcass) / (total number of daily inspections).
2.3. Statistical Analyses
All statistical analyses were performed in R version 4.3.1, with figures generated using the package ‘ggplot2’ [60]. Generalized linear models (GLMs) were performed with the glm function in the ‘lme4’ package [61], and glm.nb function in the ‘MASS’ package [62]. Linear models (LMs) were performed with lm function. We checked for the normal distribution of model residuals using simulateResiduals function in the ‘DHARMa’ package [63]. For all models, we reported likelihood ratios for the main effect and interaction between carcass-preparation investment treatments and breeding temperatures using the Anova function in the ‘car’ package [64]. Once an interaction or the treatments was found statistically significant (P < 0.05), we conducted post-hoc comparisons for the Control and the other two groups using the emmeans function with trt.vs.ctrl methods in the ‘emmeans’ package [65].
Firstly, we employed binomial (logit link) GLMs to analyze male and female care separately for each stage, to investigate the effects of carcass-preparation investment and ambient temperature on parental care during pre-hatching and post-hatching periods. Secondly, to investigate the effect of parental care on reproductive success under our carcass-preparation investment treatments and temperatures, we used GLMs with negative binomial distribution to analyse clutch size and brood size, and used LMs to analyse brood mass. Thirdly, we used LMs to analyse mean larval mass to investigate the effect of parental investment on offspring performance. Finally, we used LMs to analyse parental body mass change to investigate the effect of parental investment on their residual reproductive value. All of the models included carcass-preparation investment, ambient temperature, their interaction, and carcass size as explanatory variables. Furthermore, parental body size was included as a covariate in the models for male and female care during the pre-hatching and post-hatching periods. Female body size and pre-hatching care were incorporated into the models for clutch size, while male and female parental care were included in the models for both brood size and brood mass. Additionally, parental body size and care were considered in the models analyzing changes in parental body mass. We selected the best model for each response variable by discarding these explanatory variables that showed no significance, except for the carcass-preparation investment treatment and ambient temperature.
We conducted separate models for male and female pre- and post-hatching care, excluding their partner’s care as an explanatory variable. This approach was justified for three reasons. Firstly, our study did not aim to resolve the sexual conflict over parental care in relation to carcass-preparation investment and/or thermal conditions. Secondly, the sex-specific differences in care data distributions between pre- and post-hatching phases resulted in poor model convergence. Thirdly, incorporating higher-order interactions (e.g., carcass-preparation investment × ambient temperature × sex or carcass-preparation investment × partner’s care) introduced excessive complexity, obscuring biological interpretation.
3. Results
3.1. Effects of Carcass-Preparation Investment and Ambient Temperature on Parental Care
3.1.1. Male Pre- and Post-Hatching Care
Carcass-preparation investment, but not ambient temperature, significantly influenced male pre-hatching care (LRχ² = 7.38, df = 2, P = 0.02; LRχ² = 0.19, df = 1, P = 0.67, respectively). Males in the Reduced and Elevated groups exhibited lower levels of pre-hatching care compared to those in the Control group (Est. ± SE = -0.28 ± 0.12, z = -2.30, P = 0.04; Est. ± SE = -0.26 ± 0.12, z = -2.15, P = 0.06, respectively; Figure 2a).
For post-hatching care, the interaction between carcass-preparation investment and ambient temperature was significant (LRχ² = 10.76, df = 2, P = 0.005). Specifically, 1) under the harsh temperature (at 23°C), males exhibited reduced levels of post-hatching care across all carcass-preparation investment groups; 2) under the benign temperature (20°C), males in the Reduced group exhibited less post-hatching care compared to those in the Control group; 3) males in the Elevated and Control groups showed comparable levels of post-hatching care regardless of temperature conditions (Table 1, Figure 2b).
3.1.2. Female Pre- and Post-Hatching Care
The interaction between carcass-preparation investment and ambient temperature significantly affected female pre-hatching care (LRχ² = 6.91, df = 2, P = 0.03). Specifically, females in the Reduced and Control groups showed comparable levels of pre-hatching care under both two temperatures, whereas in the Elevated group, females breeding under the harsh temperature showed more pre-hatching care than those breeding under the benign temperature (Table 1, Figure 2c). Across all groups, females exhibited comparable levels of pre-hatching care levels within the same temperature conditions (Table 1, Figure 2c).
For female post-hatching care, carcass-preparation investment had a marginal effect (LRχ² = 5.65, df = 2, P = 0.06), while ambient temperature had a significant effect (LRχ² = 263.86, df = 1, P < 0.001). Regardless of carcass-preparation investment, females breeding under the harsh temperature exhibited reduced post-hatching care (Figure 2d).
3.2. Effects of Carcass-Preparation Investment and Ambient Temperature on Reproductive Success and Parental Body Mass Change
3.2.1. Reproductive Success
The interaction between carcass-preparation investment and ambient temperature significantly affected clutch size (LRχ² = 13.77, df = 2, P = 0.001), brood size (LRχ² = 10.94, df = 2, P = 0.004), and brood mass (F = 5.74, df = 2, P = 0.004). Specifically, under the harsh temperature, compared to the benign temperature: 1) the Reduced group produced more eggs per clutch, but smaller and lighter broods, 2) the Control group produced fewer eggs, along with smaller and lighter broods, whereas 3) the Elevated group exhibited comparable clutch size, brood size, brood mass (Table 1, 2, Figure 3a, b, c). Secondly, under the benign temperature, compared to the Control group: 1) the Reduced group produced fewer eggs, dispersed larvae of similar number, and lighter broods; 2) the Elevated group produced comparable eggs, smaller and lighter broods (Table 2, Figure 3a, b, c). Thirdly, under the harsh temperature:1) the Reduced groups produced fewer eggs than the Control group, while the Elevated group produced comparable eggs to the Control group; and 2) these groups produced broods of similar size and mass (Table 2, Figure 3a, b, c).
3.2.2. Offspring Performance
The interaction between carcass-preparation investment and ambient temperature significantly affected mean larval mass (F = 6.58, df = 2, P = 0.002). Specifically, 1) smaller dispersed larvae were produced under the harsh temperature (compared to the benign temperature) across all three groups; 2) under the benign temperature, larger dispersed larvae were produced in the Elevated group while dispersed larvae of similar mass were produced in the Reduced group, compared to the Control groups; and 3) dispersed larvae of similar mass were produced across the groups when beetle pairs bred under the harsh temperature (Table 1, Figure 3d).
3.2.3. Parental Body Mass Change
Carcass-preparation investment, ambient temperature and their interaction had no significant effect on parental body mass change for either sex (F < 2.80, P > 0.1; Figure 4). However, smaller adults gained more or lost less body mass after breeding (male: F = 25.52, P < 0.001; female: F = 23.61, P < 0.001). Additionally, male but not female body mass change was positively correlated with parental care (F = 11.14, P < 0.001).
4. Discussion
Predicting how organisms behave in response to thermal stress driven by climate change is currently one of the hotspots in the field of ecology [66]. In this study, we tested the effects of ambient temperatures and ‘past parental investment’ (i.e., parental investment in carcass preparation) on parental care strategies and reproductive success within breeding events in the burying beetles (N. vespilloides). Our results revealed that males and females exhibit distinct parental investment across pre- and post-hatching periods depending on carcass-preparation investment and ambient temperature. Consequently, the compounds of reproductive success were also significantly affected by the interaction of investment in carcass-preparation and ambient thermal conditions. Reduced investment in carcass preparation affected male parental care in both pre- and post-hatching periods, but not female parental care. In contrast, elevated carcass-preparation investment had no significant effects on parental care for either sex. Additionally, harsh temperatures negatively impacted post-hatching care in both sexes but did not influence pre-hatching care, except for females in the Elevated group.
4.1. Reduced Carcass-Preparation Investment and Ambient Temperature Impact Parental Care and Reproductive Success
Contrary to our predictions, reduced carcass-preparation investment did not lead to increased parental care in either sex. Instead, males in the Reduced group exhibited lower levels of both the pre- and post-hatching care, whereas females maintained consistent levels of care. These findings partially align with a closely related study, where beetle individuals breeding on prepared carcasses provided less parental care for their broods and abandoned them earlier [57]. Previous research in N. vespilloides, along with others in the Nicrophorus genus, has demonstrated that males but not females show greater phenotypic plasticity in caregiving behaviour in response to shifts in external cues [13,50,51,59]. This sex-specific flexibility likely explains why only males adjusted their care in response to reduced investment in carcass preparation. In the Reduced group, breeding on prepared carcasses requires less time and energy in carcass preparation, bypassing the labour-intensive tasks of carcass preparation, such as fur removal, carcass shaping, and secretion for preservation. This was in line with what we found in the male pre-hatching care. Under such conditions, males could benefit from reducing their investment in carcass preparation, while females remain under selective pressure to maintain their caregiving levels to ensure reproductive success. This pattern aligns with the “Anisogamy” theory, which proposes that sexual asymmetry in reproductive investment arises from the energetic disparity in gamete production (i.e., egg production requiring more energy than sperm) [67]. This disparity might further extend to differences in parental investment during both pre- and post-natal stages of offspring development [28]. Previous research has suggested that female burying beetles provide care at their physical limitations [14,51,68], and our findings for female pre- and post-hatching care in the Reduced and Control groups further support the idea that females bear primary responsibility for offspring care throughout the breeding period (i.e., from preparing carcasses until larval dispersal).
When considering the effects of ambient temperature, our findings, partially aligning with the prediction, revealed that both males and females decreased their post-hatching care under harsh temperatures, regardless of the carcass-preparation investment being reduced or not. However, pre-hatching care remained unchanged across temperature conditions, suggesting that beetles may have reached a baseline investment threshold for carcass preparation to avoid reproductive failure. The observed reduction in post-hatching care under high temperatures is consistent with studies showing that increased metabolic costs under thermal stress reduce subsequent parental investment [6,13,14]. As ectothermic animals, the survival and reproductive activities of insects are profoundly influenced by ambient temperature [69], with which elevated temperature increases the metabolic costs of any physical activity or behaviour, including parental care (across pre- and post-hatching periods). In this study, the consistent individual pre-hatching care observed across the two temperature conditions (both in the Reduced and Control groups) may limit male and female energy allocation to post-hatching care. Furthermore, chronic exposure to high temperatures can cause negative effects on individual physiological performance [70,71]. Such impacts align with our findings, suggesting that thermal stress may play a critical role in shaping the observed reductions in individual post-hatching care.
The interactive effect of carcass-preparation investment and thermal stress on reproductive success that we found, aligns with our findings of the adjustment of parental care behaviour. In both the Reduced and Control groups, beetle pairs breeding under harsh thermal conditions produced smaller and lighter broods with lower larval mass compared to those under benign temperature conditions, and this may be due to decreased investment in post-hatching care. This finding is consistent with previous studies on Nicrophorus species, which have documented adverse effects of elevated temperatures on reproductive outcomes [6,13,14,52,72,73]. One potential explanation is that thermal stress directly affects offspring development [70,71]. Additionally, higher temperatures may accelerate carcass decomposition, diminishing available resources for larvae [37]. This, in turn, explains why reproductive success did not differ between the Reduced and Control groups under harsh temperature conditions. This may reflect a baseline outcome driven by resource limitations affecting reproductive success.
Another interesting result is that females in the Reduced group laid fewer eggs than those in the Control group, whereas females in the Reduced group laid more eggs when they bred under the harsh temperature. In contrast, females in the Control group laid fewer eggs when they bred under the harsh temperature. These results are different from studies on N. vespilloides, which found no effect of heatwave on clutch size [6,14]. However, these findings partially align with studies on Nicrophorus nepalensis, where higher ambient temperatures had a negative effect on clutch size [72]. The observed differences in clutch size may be explained by the influence of temperature and carcass-preparation conditions on ovarian development, as ovarian development is largely dependent on carcasses [35,74]. Prepared carcasses likely contain various pheromone profiles compared to fresh carcasses, potentially influencing ovarian maturation. Elevated temperatures may amplify these pheromonal effects, but further research is needed to confirm this hypothesis.
4.2. Elevated Carcass-Preparation Investment and Ambient Temperature Impact Parental Care and Reproductive Success
Contrary to our expectations, in both sexes of beetles, increased carcass-preparation investment did not affect parental care. Individuals in the Elevated group exhibited similar parental care levels compared to those in the Control group. One possible explanation is that the additional investment in carcass preparation may improve reproductive experience, but may not impose significant energetic costs for individuals. On the one hand, beetle pairs in the Elevated group experienced ‘reproductive failure’, which may have influenced subsequent reproductive behaviour. On the other hand, these beetles may feed on the carcasses while burying them, thereby improving their body condition and offsetting their energy expenditures.
Aligning with our predictions, our results showed that, in response to high temperature, both males and females in the Elevated group decreased their post-hatching care. However, for pre-hatching care, such a response to thermal stress is different between males and females. As with previous findings, both sexes decreased parental care levels under thermal stress [6,14]. This sex-specific shift in pre-hatching care suggests that females, but not males, adjust their investment based on prior reproductive experience, such as prior investment in carcass preparation. This contradicts some previous research that suggests males exhibit greater behavioural flexibility in reproduction and parental care [13,50,51,59]. Our results indicate that females, but not males, regulate their pre-hatching care in response to environmental challenges [68].
The interactive effects of carcass-preparation investment and ambient temperature on reproductive success are complex. Firstly, under the benign thermal condition, beetles in the Elevated group produced comparable clutch sizes compared to those in the Control group, but their broods were smaller and lighter, and dispersed larvae were heavier. These results may be due to reduced female pre-hatching care leading to poorly-prepared carcasses (20℃: Control-Elevated, Est. ± SE = 0.36 ± 0.17, z = 2.17, P = 0.06, negatively impacting larval growth and survival. Additionally, prior reproductive failure may have limited female energy reserves, reducing egg viability and larval survival. Under the harsh thermal condition, there was no evidence that elevated carcass-preparation investment adversely affected reproductive success. This finding was consistent with observations of male and female pre- and post-hatching care under similar conditions, suggesting that elevated ambient temperatures limited beetles' flexibility in reproductive behaviors in response to variations in carcass-preparation investment. However, in the Elevated group, the reproductive success of beetles, excluding mean larval mass, was not adversely affected by harsh ambient thermal conditions. One possible explanation was that the increased female pre-hatching care (Elevated: 20-23 ℃, Est. ± SE = -0.43 ± 0.20, z = -2.15, P = 0.03) resulted in well-prepared carcasses, mitigating the adverse effects of heat stress on reproductive success.
Finally, relative body mass changes in males and females were similar across all treatment groups under both two thermal conditions, indicating that neither carcass-preparation investment nor temperature significantly altered energy allocation during breeding. Relative body mass change reflects individual energy allocation during breeding, in which positive values indicate greater energy conservation to secure future reproductive opportunities, and negative values suggest higher investment in current broods [57]. Therefore, our findings indicated that, as a kind of opportunist breeders, burying beetles consistently seize any available breeding opportunity, irrespective of external cues [35]. Furthermore, as shown in Figure 4, individuals in the Control group exhibited a greater difference in the relative body mass change after breeding, particularly when bred under benign temperatures. This suggests that burying beetles are more inclined to allocate energy towards current reproduction rather than conserving energy for future reproductive opportunities when exposed to external stressors, such as increased ambient temperatures, pre-prepared carcasses, and prior reproductive failure in this study.
5. Conclusion
In the present study, we investigated whether and how ambient temperature and ‘past investment’ (i.e., investment in carcass preparation) influence parental care strategies in burying beetles across pre- and post-hatching periods. Our findings showed that males and females in breeding pairs adjusted their pre- and post-hatching care differently in response to our manipulations in temperatures and carcass-preparation investments, which subsequently affected their reproductive success. Specifically, we found that (1) irrespective of ambient temperatures, males in the Reduced group decreased their pre-hatching care. (2) Across all three carcass-preparation investment groups, both males and females breeding under the harsh temperature decreased their post-hatching care. 3) Overall, the harsh temperature decreased reproductive success, and beetle pairs in the Reduced group produced fewer eggs and lighter broods, while those in the Elevated group produced smaller and lighter broods. Our findings provide new insight in exploring thermal stressor affecting organisms’ parental investment pattern, and enhance our understanding of the phenotypic plasticity in reproductive strategies that animals employ to adapt to the challenges posed by climate change. However, due to the limitations of our experimental design and manipulations, we were unable to fully eliminate the influence of mates' care when examining male and female care adjustments in response to temperature and carcass-preparation investment. For future research, we recommend more specific and accurate manipulations, such as establishing uni-female and uni-male parental care breeding events.
Author Contributions
Donghui Ma designed and conducted the experiment, analyzed the data, and wrote and revised the manuscript. Jan Komdeur and Long Ma provided guidance for the experimental design and the execution of the experiment. L.M. and J.K. edited and revised the manuscript.
Funding
This research was funded by the China Scholarship Council (CSC grant NO. 202106180024) to D.M., the Dutch Science Council grant (ALW NWO Grant No. ALWOP.531 and NWO VICI 823.01.014), and NWO TOP grant (854.11.003) to J.K.; and the Ecology Fund of the Royal Netherlands Academy of Arts and Sciences (KNAWWF/807/19021) to L.M..
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We thank the Faculty Department of Animal Care, which supplied frozen mice, laboratory space and climate rooms for this study. We appreciate the support and help with the data analyses from Dr. Maaike Versteegh. We sincerely thank Prof. Dr. Zhanqi Chen (Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences) for reading through and commenting on the earlier version of the manuscript.
Conflicts of Interest
There is no conflict of interest to declare for this study.
References
- IPCC. Climate Change 2023: Synthesis report. Contribution of working groups i, ii and iii to the sixth assessment report of the intergovernmental panel on climate change [Core Writing Team, H. Lee and J. Romero (Eds.)]. IPCC, Geneva, Switzerland, 2023.
- Saunders, D.A.; Mawson, P.; Dawson, R. The impact of two extreme weather events and other causes of death on Carnaby’s black cockatoo: A promise of things to come for a threatened species? Pac. Conserv. Biol. 2011, 17, 141–148. [Google Scholar] [CrossRef]
- Fragueira, R.; Helfenstein, F.; Fischer, K.; Beaulieu, M. Birds of different morphs use slightly different strategies to achieve similar reproductive performance following heatwave exposure. J. Anim. Ecol. 2021, 90, 2594–2608. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.; Simmons, L.W. Behavioural dynamics of biparental care in the dung beetle Onthophagus taurus. Anim. Behav. 2002, 64, 65–75. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Bressac, C.; Chevrier, C. Heat stress affects male reproduction in a parasitoid wasp. J. Insect Physiol. 2013, 59, 248–254. [Google Scholar] [CrossRef]
- Pilakouta, N.; Allan, D.; Moore, E.; Russell, A.A. chronic and acute thermal stressors have non-additive effects on fertility. Proc. Royal Soc. B 2024, 291, rspb20241086. [Google Scholar] [CrossRef]
- Sales, K.; Vasudeva, R.; Dickinson, M.E.; Godwin, J.L.; Lumley, A.J.; Michalczyk, Ł.; Hebberecht, L.; Thomas, P.; Franco, A.; Gage, M.J.G. Experimental heatwaves compromise sperm function and cause transgenerational damage in a model insect. Nat. Commun. 2018, 9, 4771. [Google Scholar] [CrossRef]
- Chevrier, C.; Nguyen, T.M.; Bressac, C. Heat shock sensitivity of adult male fertility in the parasitoid wasp Anisopteromalus calandrae (Hymenoptera, Pteromalidae). J. Therm. Biol. 2019, 85, 102419. [Google Scholar] [CrossRef]
- Porcelli, D.; Gaston, K.J.; Butlin, R.K.; Snook, R.R. Local adaptation of reproductive performance during thermal stress. J. Evol. Biol. 2017, 30, 422–429. [Google Scholar] [CrossRef]
- Martinet, B.; Zambra, E.; Przybyla, K.; Lecocq, T.; Anselmo, A.; Nonclercq, D.; Rasmont, P.; Michez, D.; Hennebert, E. Mating under climate change: impact of simulated heatwaves on the reproduction of model pollinators. Funct. Ecol. 2021, 35, 739–752. [Google Scholar] [CrossRef]
- Rukke, B.A.; Sivasubramaniam, R.; Birkemoe, T.; Aak, A. Temperature stress deteriorates bed bug (Cimex lectularius) populations through decreased survival, fecundity and offspring success. PLOS ONE 2018, 13, e0193788. [Google Scholar] [CrossRef]
- McCowan, L.S.C.; Griffith, S.C. Baked Eggs: Catastrophic heatwave-induced reproductive failure in the desert-adapted Zebra finch (Taeniopygia guttata). Ibis 2021, 163, 1207–1216. [Google Scholar] [CrossRef]
- Moss, J.B.; Moore, A.J. Constrained flexibility of parental cooperation limits adaptive responses to harsh conditions. Evol. 2021, 75, 1835–1849. [Google Scholar] [CrossRef]
- Pilakouta, N.; Sellers, L.; Barratt, R.; Ligonniere, A. The consequences of heatwaves for animal reproduction are timing-dependent. Funct. Ecol. 2023, 1–9. [Google Scholar] [CrossRef]
- Wiley, E.M.; Ridley, A.R. The effects of temperature on offspring provisioning in a cooperative breeder. Anim. Behav. 2016, 117, 187–195. [Google Scholar] [CrossRef]
- Kumar, S.; Kler, T.K.; Sekhon, G.S.; Sahni, T. Impacts on avian migratory patterns due to climate change and hormonal disruption: a review. Mitig. Adapt. Strateg. Glob. Change 2024, 29, 69. [Google Scholar] [CrossRef]
- Both, C.; Bouwhuis, S.; Lessells, C.M.; Visser, M.E. Climate change and population declines in a long-distance migratory bird. Nature 2006, 441, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Charmantier, A.; McCleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef]
- Dawson, R.D.; Lawrie, C.C.; O’Brien, E.L. The importance of microclimate variation in determining size, growth and survival of avian offspring: experimental evidence from a cavity nesting passerine. Oecologia 2005, 144, 499–507. [Google Scholar] [CrossRef]
- McKechnie, A.E. Physiological and morphological effects of climate change. In Effects of Climate Change on Birds; 2nd ed.; Dunn, P.O., Møller, A.P., Eds, Oxford University Press: Oxford, UK, 2019; pp. 201–133. [Google Scholar]
- Sun, B.-J.; Wang, Y.; Wang, Y.; Lu, H.-L.; Du, W.-G. Anticipatory parental effects in a subtropical lizard in response to experimental warming. Front. Zool. 2018, 15, 51. [Google Scholar] [CrossRef]
- Sidhu, K.K.; Zafeiri, S.; Malcolm, C.; Caplat, P.; Lancaster, L.T.; Bocedi, G.; Pilakouta, N. Heatwaves during early development have long-term consequences for parental care in adulthood. Anim. Behav. 2024, 217, 65–72. [Google Scholar] [CrossRef]
- Walsh, B.S.; Parratt, S.R.; Hoffman, A.A.; Atkinson, D.; Snook, R.R.; Bretman, A.; Price, T.A.R. The impact of climate change on fertility. Trends Ecol. Evol. 2019, 34, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. Trade-offs in life-history evolution. Funct. Ecol. 1989, 3, 259–268. [Google Scholar] [CrossRef]
- Gonzalez, O.; Zedrosser, A.; Pelletier, F.; Swenson, J.E.; Festa-Bianchet, M. Litter reductions reveal a trade-off between offspring size and number in Brown bears. Behav. Ecol. Sociobiol. 2012, 66, 1025–1032. [Google Scholar] [CrossRef]
- Richardson, J.; Stephens, J.; Smiseth, P.T. Increased allocation to reproduction reduces future competitive ability in a burying beetle. J. Anim. Ecol. 2020, 89, 1918–1926. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.S.A.; Nakagawa, S. The costs of parental care: a meta-analysis of the trade-off between parental effort and survival in birds. J. Evol. Biol. 2012, 25, 1911–1917. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R. Parental investment and sexual selection. In sexual selection and the descent of man; Campbell, B., Eds, Aldine: Chicago, US, 1972; pp. 1871–1971. [Google Scholar]
- Coleman, R.M.; Gross, M.R. Parental investment theory: the role of past investment. Trends Ecol. Evol. 1991, 6, 404–406. [Google Scholar] [CrossRef]
- Bonnet, X.; Lourdais, O.; Shine, R.; Naulleau, G. Reproduction in a typical capital breeder: costs, currencies, and complications in the Aspic viper. Ecol. 2002, 83, 2124–2135. [Google Scholar] [CrossRef]
- van Noordwijk, A.J.; de Jong, G. Acquisition and allocation of resources: their influence on variation in life history tactics. Am. Nat. 1986, 128, 137–142. [Google Scholar] [CrossRef]
- Glazier, D.S. Is fatter fitter? Body storage and reproduction in ten populations of the freshwater amphipod Gammarus minus. Oecologia 2000, 122, 335–345. [Google Scholar] [CrossRef]
- Messina, F.J.; Fry, J.D. Environment-dependent reversal of a life history trade-off in the seed beetle Callosobruchus maculatus. J. Evol. Biol. 2003, 16, 501–509. [Google Scholar] [CrossRef]
- Angilletta, M.J. Thermal adaptation: a theoretical and empirical synthesis. Oxford University Press: Oxford, UK, 2009; pp.88–125.
- Scott, M.P. The ecology and behavior of burying beetles. Annu. Rev. Entomol. 1998, 43, 595–618. [Google Scholar] [CrossRef]
- Bartlett, J. The behavioural ecology of the burying beetle. Degree of Doctor of Philosophy, University of Edinburgh, Edinburgh, 1987.
- Arce, A.N.; Johnston, P.R.; Smiseth, P.T.; Rozen, D.E. Mechanisms and fitness effects of antibacterial defences in a carrion beetle. J. Evol. Biol. 2012, 25, 930–937. [Google Scholar] [CrossRef]
- Müller, J.K.; Eggert, A.-K.; Sakaluk, S.K. Carcass maintenance and biparental brood care in burying beetles: are males redundant? Ecol. Entomol. 1998, 23, 195–200. [Google Scholar] [CrossRef]
- Eggert, A.-K.; Reinking, M.; Müller, J.K. Parental care improves offspring survival and growth in burying beetles. Anim. Behav. 1998, 55, 97–107. [Google Scholar] [CrossRef]
- Schrader, M.; Hughes, P.; Jenkins, S.; Kusher, I.; Lopez, J.; Oglesby, H.; McGhee, K.E. Can age-related changes in parental care modulate inbreeding depression? A test using the burying beetle, Nicrophorus orbicollis. Ecol. Evol. 2022, 12, e9391. [Google Scholar] [CrossRef]
- Pilakouta, N.; Jamieson, S.; Moorad, J.A.; Smiseth, P.T. Parental care buffers against inbreeding depression in burying beetles. PNAS 2015, 112, 8031–8035. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.; Ashworth, C.M. Brood size and fitness in Nicrophorus Vespilloides (Coleoptera: Silphidae). Behav. Ecol. Sociobiol. 1988, 22, 429–434. [Google Scholar] [CrossRef]
- Keppner, E.M.; Laubenthal, M.; Prang, M.A.; Conrad, T.; Steiger, S. Harsh nutritional environment has positive and negative consequences for family living in a burying beetle. Ecol. Evol. 2023, 13, e9699. [Google Scholar] [CrossRef]
- Richardson, J.; Smiseth, P.T. Nutrition during sexual maturation and at the time of mating affects mating behaviour in both sexes of a burying beetle. Anim. Behav. 2019, 151, 77–85. [Google Scholar] [CrossRef]
- Richardson, J.; Smiseth, P.T. Effects of variation in resource acquisition during different stages of the life cycle on life-history traits and trade-offs in a burying beetle. J. Evol. Biol. 2019, 32, 19–30. [Google Scholar] [CrossRef]
- Eggert, A.-K.; Müller, J.K. Timing of oviposition and reproductive skew in cobreeding female burying beetles (Nicrophorus vespilloides). Behav. Ecol. 2000, 11, 357–366. [Google Scholar] [CrossRef]
- Komdeur, J.; Schrama, M.J.J.; Meijer, K.; Moore, A.J.; Beukeboom, L.W. Cobreeding in the burying beetle, Nicrophorus vespilloides: tolerance rather than cooperation. Ethol. 2013, 119, 1138–1148. [Google Scholar] [CrossRef]
- Ma, L.; Versteegh, M.A.; Hammers, M.; Komdeur, J. Sex-specific influence of communal breeding experience on parenting performance and fitness in a burying beetle. R. Soc. Open Sci. 2022, 9, 211179. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.; Smiseth, P.T. Maternity uncertainty in cobreeding beetles: females lay more and larger eggs and provide less care. Behav. Ecol. 2020, 31, 641–650. [Google Scholar] [CrossRef]
- Smiseth, P.T.; Dawson, C.; Varley, E.; Moore, A.J. How do caring parents respond to mate loss? differential response by males and females. Anim. Behav. 2005, 69, 551–559. [Google Scholar] [CrossRef]
- Wang, W.; Ma, L.; Versteegh, M.A.; Wu, H.; Komdeur, J. Parental care system and brood size drive sex difference in reproductive allocation: an experimental study on burying beetles. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Grew, R.; Ratz, T.; Richardson, J.; Smiseth, P. Parental care buffers against effects of ambient temperature on offspring performance in an insect. Behav. Ecol. 2019, 30, 1443–1450. [Google Scholar] [CrossRef]
- De Gasperin, O.; Kilner, R.M. Interspecific interactions change the outcome of sexual conflict over prehatching parental investment in the burying beetle Nicrophorus vespilloides. Ecol. Evol. 2015, 5, 5552–5560. [Google Scholar] [CrossRef]
- De Gasperin, O.; Duarte, A.; Troscianko, J.; Kilner, R.M. Fitness costs associated with building and maintaining the burying beetle’s carrion nest. Sci. Rep. 2016, 6, 1–6. [Google Scholar] [CrossRef]
- Carter, D.O.; Yellowlees, D.; Tibbett, M. Temperature affects microbial decomposition of cadavers (Rattus rattus) in contrasting soils. Appl. Soil Ecol. 2008, 40, 129–137. [Google Scholar] [CrossRef]
- Mattey, S.N.; Strutt, L.; Smiseth, P.T. Intergenerational effects of inbreeding in Nicrophorus vespilloides: offspring suffer fitness costs when either they or their parents are inbred. J. Evol. Biol. 2013, 26, 843–853. [Google Scholar] [CrossRef]
- Wang, W.; Ma, L.; Versteegh, M.A.; Wu, H.; Komdeur, J. Detection of reproductive trade-offs is influenced by resource availability and maintenance: an experimental study in the burying beetle (Nicrophorus vespilloides). Behav. Ecol. Sociobiol. 2022, 76. [Google Scholar] [CrossRef]
- Bladon, E.K.; English, S.; Pascoal, S.; Kilner, R.M. Early-life effects on body size in each sex interact to determine reproductive success in the burying beetle Nicrophorus vespilloides. J. Evol. Biol. 2020, 33, 1725–1734. [Google Scholar] [CrossRef]
- Jarrett, B.J.M.; Schrader, M.; Rebar, D.; Houslay, T.M.; Kilner, R.M. Cooperative interactions within the family enhance the capacity for evolutionary change in body size. Nat. Ecol. Evol. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant graphics for data analysis. Springer: New York, US, 2016.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern applied statistics with S, 3rd ed.; Springer: New York, US, 2002. [Google Scholar]
- Hartig, F. Residual diagnostics for hierarchical (Multi-Level / Mixed) regression models [R package DHARMa version 0.4.6], 2022.
- Fox, J.; Weisberg, S.; Price, B. An R Companion to Applied Regression, 3rd ed.; Sage: Los Angeles, US, 2019.
- Lenth, R.V. emmeans: Estimated marginal means, aka least-squares means. R package version 1.7.1. R Foundation for Statistical Computing, 2021.
- Briscoe, N.J.; Morris, S.D.; Mathewson, P.D.; Buckley, L.B.; Jusup, M.; Levy, O.; Maclean, I.M.D.; Pincebourde, S.; Riddell, E.A.; Roberts, J.A.; et al. Mechanistic forecasts of species responses to climate change: the promise of biophysical ecology. Glob. Chang. Biol. 2023, 29, 1451–1470. [Google Scholar] [CrossRef]
- Lehtonen, J.; Kokko, H.; Parker, G.A. What do isogamous organisms teach us about sex and the two sexes? Philos. Trans. R. Soc. Lond. B, Biol. Sci. 2016, 371, 20150532. [Google Scholar] [CrossRef]
- Cotter, S.C.; Kilner, R.M. Sexual division of antibacterial resource defence in breeding burying beetles, Nicrophorus vespilloides. J. Anim. Ecol. 2010, 79, 35–43. [Google Scholar] [CrossRef]
- Abram, P.K.; Boivin, G.; Moiroux, J.; Brodeur, J. Behavioural effects of temperature on ectothermic animals: unifying thermal physiology and behavioural plasticity. Biol. Rev. 2017, 92, 1859–1876. [Google Scholar] [CrossRef]
- Evans, R.K.; Toews, M.D.; Sial, A.A. Impact of short- and long-term heat stress on reproductive potential of drosophila Suzukii matsumura (Diptera: Drosophilidae). J. Therm. Biol. 2018, 78, 92–99. [Google Scholar] [CrossRef]
- Dowd, W.W.; King, F.A.; Denny, M.W. Thermal Variation, Thermal extremes and the physiological performance of individuals. J. Therm. Biol. 2015, 218, 1956–1967. [Google Scholar] [CrossRef]
- Malik, T.G.; Jarrett, B.J.M.; Sun, S.-J. The effect of experimental warming on reproductive performance and parental care in the burying beetle Nicrophorus nepalensis. R. Soc. Open Sci. 2024, 11, 240653. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.L.; Howard, D.R.; Hall, C.L. The thermal ecology of burying beetles: temperature influences reproduction and daily activity in Nicrophorus marginatus. Ecol. Entomol. 2021, 46, 1266–1272. [Google Scholar] [CrossRef]
- Pellissier Scott, M.; Traniello, J.F.A. Behavioural Cues Trigger ovarian development in the burying beetle, Nicrophorus tomentosus. J. Insect Physiol. 1987, 33, 693–696. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of the experiment. On the left, three levels of investment in carcass preparation are shown: Reduced (virgin beetle pair bury one already prepared mouse carcass by another beetle pair and breed on it), Control (virgin beetle pair bury one newly thawed dead mouse carcass and breed on it), and Elevated (beetle pair that experienced carcass preparation once bury one newly thawed dead mouse carcass and breed on it). On the right, two ambient temperatures indicate the conditions for carcass preparation and breeding events. The beetle pairs without colouration (e.g., pair in the Reduced group) do not engage in carcass preparation, while those with blue colouration prepare a carcass once, and those with orange colouration carry out the task twice.
Figure 1.
Schematic diagram of the experiment. On the left, three levels of investment in carcass preparation are shown: Reduced (virgin beetle pair bury one already prepared mouse carcass by another beetle pair and breed on it), Control (virgin beetle pair bury one newly thawed dead mouse carcass and breed on it), and Elevated (beetle pair that experienced carcass preparation once bury one newly thawed dead mouse carcass and breed on it). On the right, two ambient temperatures indicate the conditions for carcass preparation and breeding events. The beetle pairs without colouration (e.g., pair in the Reduced group) do not engage in carcass preparation, while those with blue colouration prepare a carcass once, and those with orange colouration carry out the task twice.
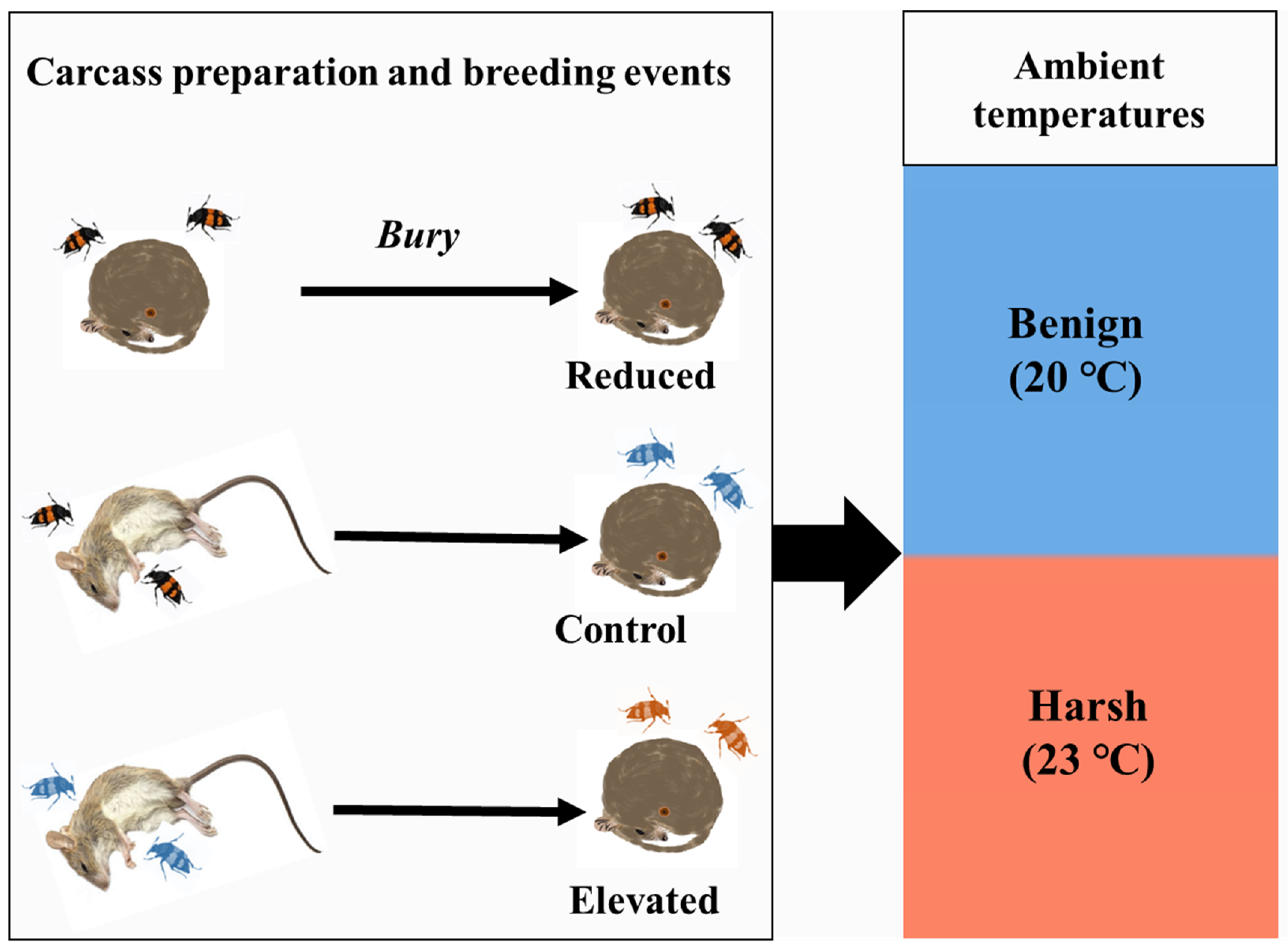
Figure 2.
Effect of carcass-preparation treatments and ambient temperatures on male (a) pre-hatching and (b) post-hatching parental care, and female (c) pre-hatching and (d) post-hatching parental care. Boxplots show median, interquartile range, and minimum/maximum range. The coloured dots represent the real values of individual post-hatching care, while the black dots represent outliers. The letters in (a-c) indicate significant trt.vs.ctrl comparisons from the post-hoc analysis, with which the same letter represents no difference. In (a), only the differences among these three treatments of carcass-preparation investment (i.e., Reduced vs. Control, Elevated vs. Control) were compared as the interaction of fixed factors did not show significance.
Figure 2.
Effect of carcass-preparation treatments and ambient temperatures on male (a) pre-hatching and (b) post-hatching parental care, and female (c) pre-hatching and (d) post-hatching parental care. Boxplots show median, interquartile range, and minimum/maximum range. The coloured dots represent the real values of individual post-hatching care, while the black dots represent outliers. The letters in (a-c) indicate significant trt.vs.ctrl comparisons from the post-hoc analysis, with which the same letter represents no difference. In (a), only the differences among these three treatments of carcass-preparation investment (i.e., Reduced vs. Control, Elevated vs. Control) were compared as the interaction of fixed factors did not show significance.
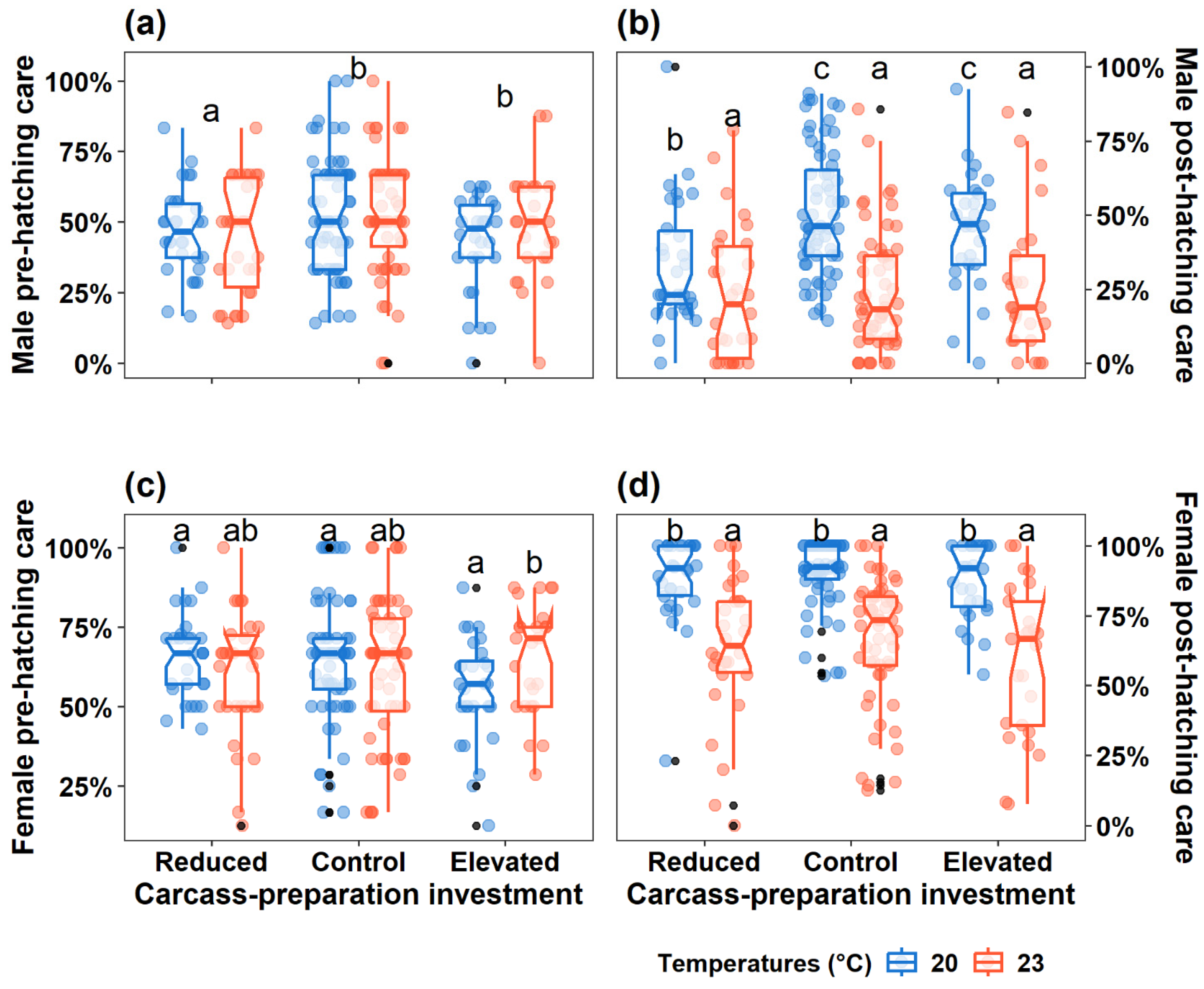
Figure 3.
Effect of carcass-preparation treatment and ambient temperatures on (a) clutch size, (b) brood size, (c) brood mass, and (d) mean larval mass. Boxplots show median, interquartile range, and minimum/maximum range. The coloured dots represent the real values, while the black dots represent each treatment's outliers. The letters indicate significant trt.vs.ctrl comparisons from the post-hoc analysis, with which the same letter represents no difference.
Figure 3.
Effect of carcass-preparation treatment and ambient temperatures on (a) clutch size, (b) brood size, (c) brood mass, and (d) mean larval mass. Boxplots show median, interquartile range, and minimum/maximum range. The coloured dots represent the real values, while the black dots represent each treatment's outliers. The letters indicate significant trt.vs.ctrl comparisons from the post-hoc analysis, with which the same letter represents no difference.
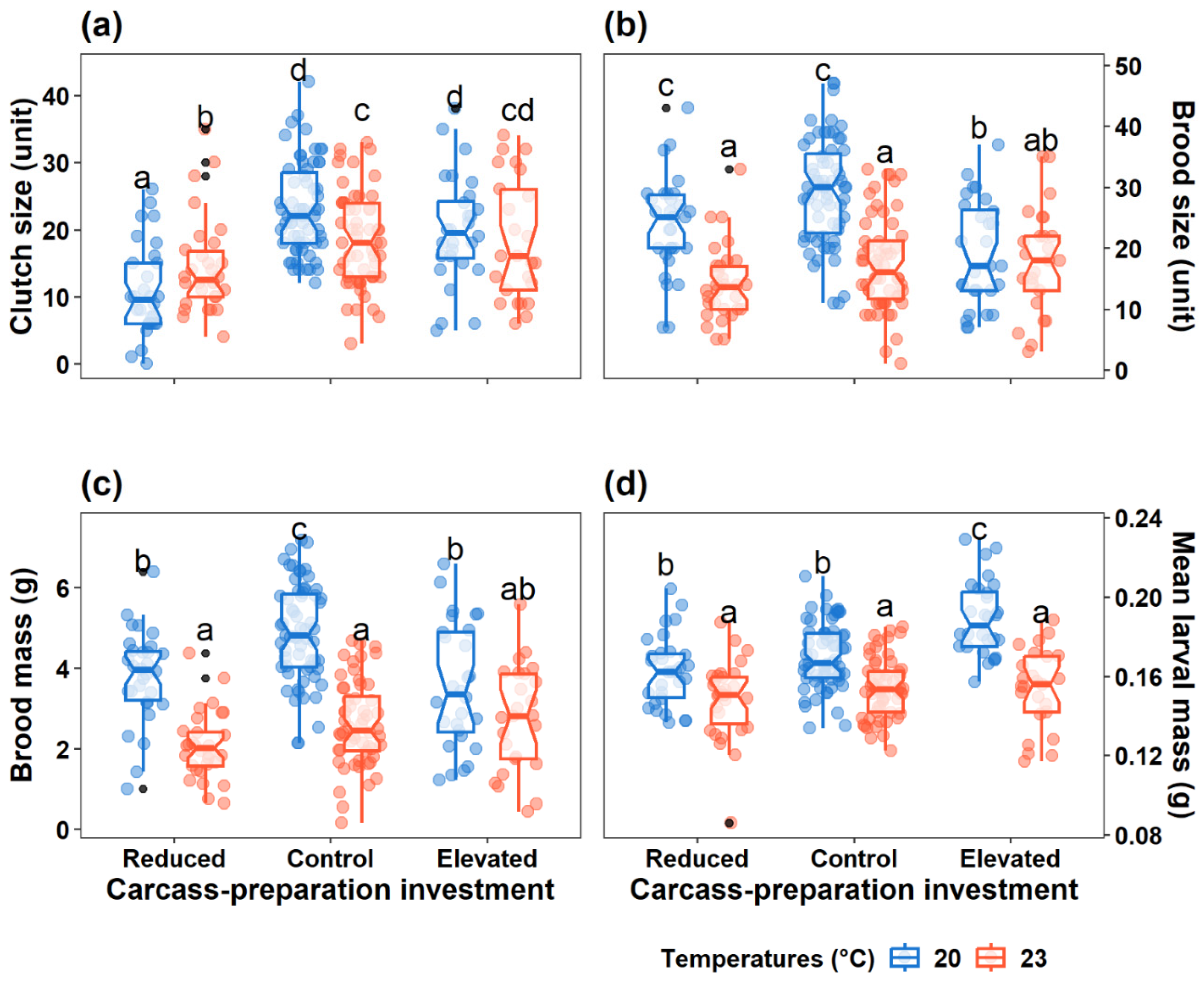
Figure 4.
Effect of carcass-preparation treatment and ambient temperature on (a) male and (b) female body mass change during breeding. The coloured dots represent the values of relative parental body mass change. Mean values are illustrated as dark-coloured dots with error bars. The red dashed line (y = 0) represents no body mass change during breeding.
Figure 4.
Effect of carcass-preparation treatment and ambient temperature on (a) male and (b) female body mass change during breeding. The coloured dots represent the values of relative parental body mass change. Mean values are illustrated as dark-coloured dots with error bars. The red dashed line (y = 0) represents no body mass change during breeding.
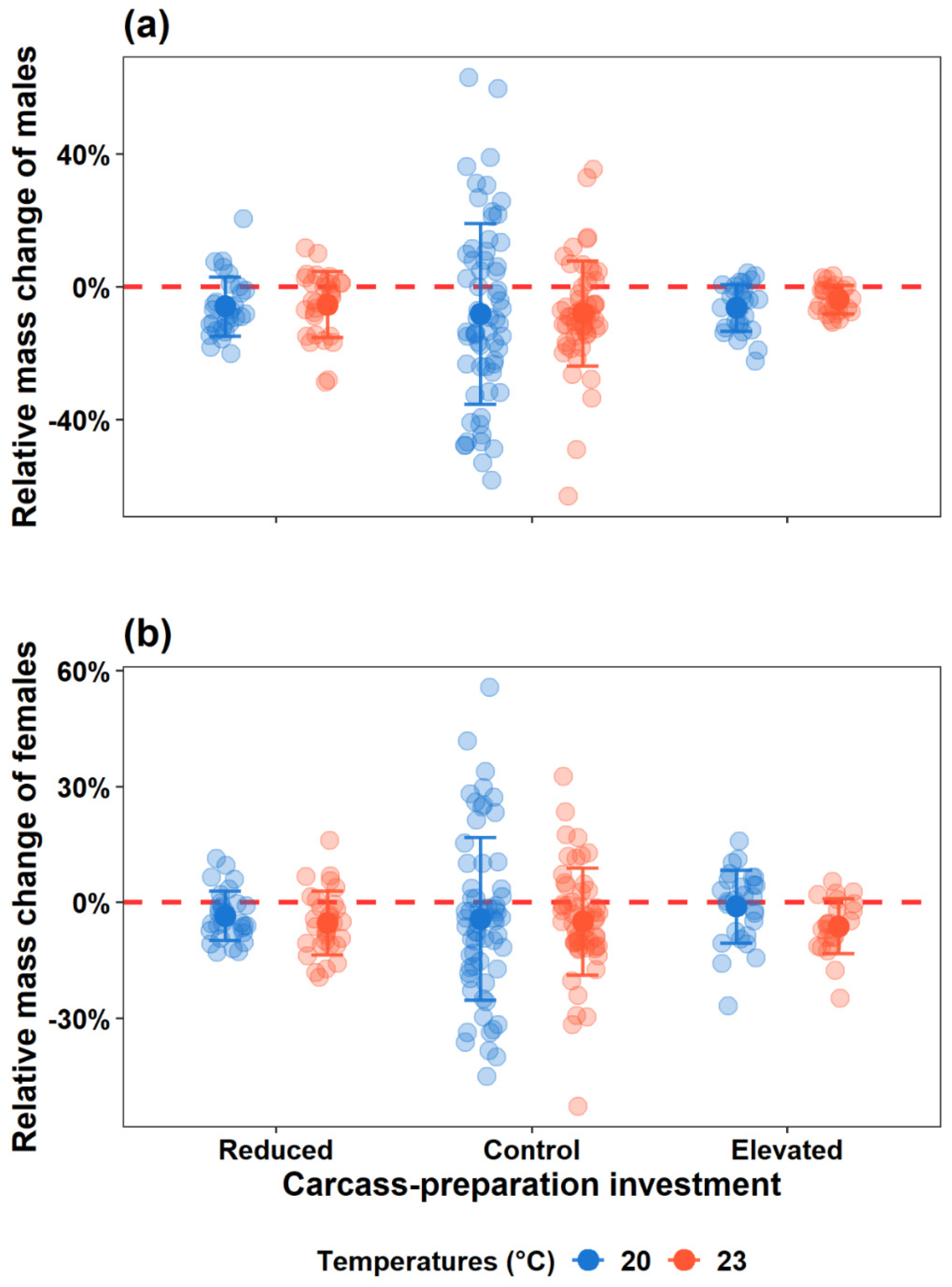

Table 1.
The trt.vs.ctrl comparison results from the post-hoc test for GLMs on male post-hatching care, female pre-hatching care, and offspring performance (i.e., mean larval mass).
Table 1.
The trt.vs.ctrl comparison results from the post-hoc test for GLMs on male post-hatching care, female pre-hatching care, and offspring performance (i.e., mean larval mass).
| Variables | Male post-hatching care | Female pre-hatching care | Mean larval mass | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Est.± SE | z | P | Est.± SE | z | P | Est.± SE | t | P | |
| Control: 20-23 ℃ | 1.13± 0.11 | 9.84 | < 0.001 | 0.14± 0.15 | 0.97 | 0.33 | 0.02± 0.003 | 4.50 | < 0.001 |
| Reduced: 20-23 ℃ | 0.48± 0.16 | 2.95 | 0.003 | 0.23± 0.20 | 1.15 | 0.25 | 0.01± 0.005 | 3.15 | 0.002 |
| Elevated: 20-23 ℃ | 0.91± 0.17 | 5.52 | < 0.001 | -0.43± 0.20 | -2.15 | 0.03 | 0.04± 0.005 | 7.32 | < 0.001 |
| 20℃: Control-Reduced | 0.71± 0.13 | 5.33 | < 0.001 | -0.05±0.17 | -0.31 | 0.94 | 0.008± 0.004 | 1.94 | 0.11 |
| 20℃: Control-Elevated | 0.21± 0.13 | 1.69 | 0.17 | 0.36±0.17 | 2.17 | 0.06 | -0.019± 0.004 | -4.71 | < 0.001 |
| 23℃: Control-Reduced | 0.06± 0.15 | 0.41 | 0.90 | 0.03± 0.18 | 0.19 | 0.98 | 0.006± 0.004 | 1.43 | 0.28 |
| 23℃: Control-Elevated | -0.002± 0.16 | -0.01 | 1.00 | -0.21±0.19 | -1.15 | 0.44 | 0.0005± 0.004 | 0.11 | 0.99 |
Note: Est. is the abbreviation of estimate. SE represents standard error.
Table 2.
The trt.vs.ctrl comparison results from the post-hoc test for GLMs on the components of reproductive success (clutch size, brood size, and brood mass).
Table 2.
The trt.vs.ctrl comparison results from the post-hoc test for GLMs on the components of reproductive success (clutch size, brood size, and brood mass).
| Variables | Clutch size | Brood size | Brood mass | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Est.± SE | z | P | Est.± SE | z | P | Est.± SE | t | P | |
| Control: 20-23 ℃ | 0.22± 0.07 | 2.91 | 0.004 | 0.23± 0.08 | 2.87 | 0.004 | 1.41± 0.24 | 5.89 | < 0.001 |
| Reduced: 20-23 ℃ | -0.29± 0.11 | -2.53 | 0.01 | 0.26± 0.10 | 2.52 | 0.01 | 1.00± 0.30 | 3.37 | 0.001 |
| Elevated: 20-23 ℃ | 0.08± 0.11 | 0.71 | 0.48 | -0.14± 0.11 | -1.34 | 0.19 | 0.20± 0.31 | 0.63 | 0.53 |
| 20℃: Control-Reduced | 0.76± 0.10 | 7.80 | < 0.001 | 0.08± 0.08 | 0.93 | 0.58 | 0.61± 0.25 | 2.41 | 0.03 |
| 20℃: Control-Elevated | 0.15± 0.09 | 1.59 | 0.21 | 0.32± 0.08 | 3.90 | < 0.001 | 1.03± 0.25 | 4.12 | < 0.001 |
| 23℃: Control-Reduced | 0.25± 0.10 | 2.63 | 0.02 | 0.11± 0.09 | 1.16 | 0.43 | 0.20± 0.26 | 0.79 | 0.68 |
| 23℃: Control-Elevated | 0.009± 0.10 | 0.09 | 0.99 | -0.05± 0.09 | -0.54 | 0.83 | -0.19± 0.26 | -0.72 | 0.72 |
Note: Est. is the abbreviation of estimate. SE represents standard error.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



